PLC-ε: A Shared Effector Protein in Ras-, Rho-, and Gαβγ-Mediated Signaling
- Address correspondence to TKH. E-mail tkh{at}med.unc.edu; fax 919-966-5640.
Abstract
The conceptual segregation of G protein–stimulated cell signaling responses into those mediated by heterotrimeric G proteins versus those promoted by small GTPases of the Ras superfamily is no longer vogue. PLC-ε , an isozyme of the phospholipase C (PLC) family, has been identified recently and dramatically extends our understanding of the crosstalk that occurs between heterotrimeric and small monomeric GTPases. Like the widely studied PLC-β isozymes, PLC-ε is activated by Gβγ released upon activation of heterotrimeric G proteins. However, PLC-ε markedly differs from the PLC-β isozymes in its capacity for activation by Gα12/13 - but not Gαq -coupled receptors. PLC-ε contains two Ras-associating domains located near the C terminus, and H-Ras regulates PLC-ε as a downstream effector. Rho also activates PLC-ε , but in a mechanism independent of the C-terminal Ras-associating domains. Therefore, Ca2+ mobilization and activation of protein kinase C are signaling responses associated with activation of both H-Ras and Rho. A guanine nucleotide exchange domain conserved in the N terminus of PLC-ε potentially confers a capacity for activators of this isozyme to cast signals into additional signaling pathways mediated by GTPases of the Ras superfamily. Thus, PLC-ε is a multifunctional nexus protein that senses and mediates crosstalk between heterotrimeric and small GTPase signaling pathways.
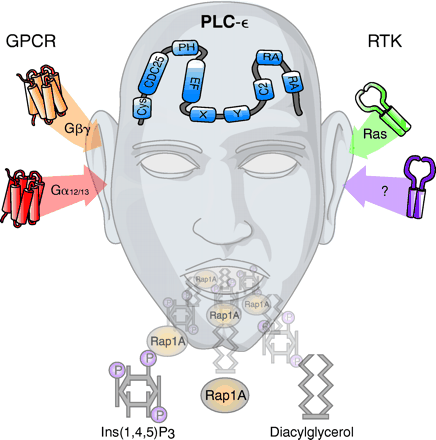
Introduction
Two decades ago, Robert Michell (1, 2) , Michael Berridge (3) , and their colleagues discovered the role of phospholipase C (PLC) in initiating the intracellular events elicited by many hormones, neurotransmitters, and growth factors. Delineation of the enzymatic hydrolysis of phosphatidylinositol 4,5-bisphosphate [PtdIns(4,5)P2 ] by PLC as the first of many biochemical steps underlying both the mobilization of intracellular Ca2+ and activation of protein kinase C (Figure 1⇓) paved the way for understanding signaling pathways that emanate from activation of other lipid-metabolizing enzymes (e.g., phospholipase A2 , phospholipase D, and phosphoinositol 3-kinase).
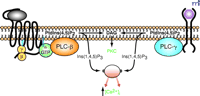
Receptor-promoted activation of phospholipase C and generation of second messengers. A prototypical G protein–coupled receptor is schematized as a black string (left) with seven transmembrane domains. Binding of a hormone ligand (gray oval) results in activation and dissociation of heterotrimeric G proteins into free Gβγ and GTP-bound α subunits. In accordance with the historical view, the GTP-bound α subunit activates an isoform of PLC-β, which hydrolyzes phosphatidylinositol 4,5-bisphosphate [PtdIns(4,5)P2] into inositol 1,4,5-trisphosphate [Ins(1,4,5)P3], which raises the cytosolic concentration of calcium ions ([Ca2+]i) and diacylglycerol (DAG), an activator of protein kinase C (PKC). Also schematized is a receptor tyrosine kinase (right) bound by an extracellular growth factor (pink circle); the epidermal growth factor receptor (EGFR) conforms to this schematization. Traditionally, this second class of receptor was proposed to initiate an inositol lipid response through activation of an isoform of PLC-γ as a consequence of tyrosine phosphorylation of the isozyme.
Isozymes of PLC (e.g., PLC-β , -γ , and -δ ) were identified over a decade ago (4) , and a subset of these was found to be regulated by heterotrimeric G proteins (5–8) coupled to cell surface receptors, or G protein–coupled receptors (GPCRs). Specifically, the PLC-β isozymes (PLC-β 1, -β 2, -β 3, and -β 4) are activated by Gα subunits of the Gq family of heterotrimeric G proteins; PLC-β 2 and PLC-β 3 are also activated by Gβγ (9–11) . PLC-γ isozymes contain Src homology 2 and 3 (SH2 and SH3) domains and are thus regulated by tyrosine kinases and membrane translocation (12–14) . The importance, if any, of PLC-δ in receptor-promoted inositol lipid signaling remains largely undefined (4) . Another recently identified isozyme, PLC-ζ , with distribution apparently limited to sperm, is reported to trigger the Ca2+ oscillations that occur in eggs upon in vitro fertilization (15, 16) . The mechanism of regulation of PLC-ζ is unknown.
Our conception of G protein–mediated regulation of inositol lipid signaling has been dramatically altered by the recent discovery of a new PLC isozyme, PLC-ε (17–20) . This G protein effector is dually regulated upstream by small monomeric GTPases of the Ras (18, 20) and Rho (21) families and by heterotrimeric G proteins (19, 22) . Moreover, downstream signal(s) in addition to the mobilization of Ca2+ and activation of protein kinase C emanate from the activity of PLC-ε as a guanine nucleotide exchange factor (GEF) for GTPases of the Ras superfamily (18, 19, 23, 24) . Thus, upstream signals from small monomeric GTPases as well as those from heterotrimeric G proteins converge on PLC-ε , and multiple downstream signals emanate from PLC-ε . We discuss here the current understanding of this pivotal signaling protein.
Identification of PLC-ε
Kataoka and his colleagues (17) identified a C. elegans protein, PLC210, in a yeast two-hybrid screen to identify proteins that bind LET-60 (i.e., a C. elegans Ras ortholog). This 210-kDa (1898-residue) protein possesses sequences that are homologous to the catalytic domain (i.e., the X and Y boxes) and the Ca2+ /lipid-binding domain (i.e., the C2 domain) that typify all characterized PLCs. Moreover, the unique presence of a C-terminal Ras-associating (RA) domain and an N-terminal Ras GEF domain suggested a bifunctional regulatory potential for PLC-ε as both an effector of Ras and an activator of downstream Ras GTPase signaling pathways.
Three laboratories independently applied sequence information from existing expressed sequence tag databases to clone mammalian orthologs of PLC210, designated PLC-ε (18–20) . Two human splice variants of 1994 and 2303 residues, which differ only in their N termini, and a single rat isoform of 2281 residues were reported. Further examination of databases suggests the existence of additional splice variants of human PLC-ε . The mammalian isoforms exhibit a domain organization similar to the C. elegans ortholog (Figure 2⇓). The functionally characterized domains of PLC-ε are considered below.
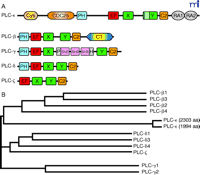
Domain organization and amino acid sequence comparisons among human PLC isozymes.
A. The five classes of phospholipase C isoforms are distinguished by differing structural domains, drawn here to relative scale in the full-length isozymes. The unique 60–70-residue “insert” is identified in the PLC-ε Y box.
B. A dendogram of the five groups of PLC isoforms is presented. The classification of PLC isoforms into five groups is reflected at the overall sequence level.
(Domain identification: Cys, cysteine-rich region; CDC25, guanine nucleotide exchange domain conserved among guanine nucleotide exchange factors (GEF); PH, pleckstrin homology domain; EF, EF-hand domain; X and Y, core catalytic domains (i.e., a TIM barrel); C2, Ca2+/lipid-binding domain; RA1 and RA2, Ras-associating domains. Genbank protein accession numbers for the known human PLC isozymes are as follows: PLC-ε, [1994 aa] AAG28341, [2303 aa] AAG17145; PLC-β1, Q9NQ66; PLC-β2, Q00722; PLC-β3, Q01970; PLC-β4, Q15147; PLC-γ1, P19174; PLC-γ2, P16885; PLC-δ1, P51178; PLC-δ3, NP_588614; PLC-δ4, NP_116115; PLC-ζ, NP_149114.)
Ras-Associating Domains
RA domains, sequences of approximately 100 residues found in many proteins, interact directly with Ras-family GTPases (25) . Although RA domains share low sequence homology with the conserved sequences known as Ras-binding domains, crystal structures have revealed that these two Ras-interacting domains share similar three-dimensional folds (26, 27) . PLC-ε contains two consecutive RA domains (RA1 and RA2) near its C terminus. Kataoka and colleagues initially demonstrated GTP-dependent binding of H-Ras to the RA domain of PLC210 in vitro (17) and subsequently reported interaction of both H-Ras and members of the Rap subfamily of GTPases to the RA2 domain of human PLC-ε (18, 24) . Kelley and coworkers (20) confirmed not only that the RA2 domain of rat PLC-ε binds H-Ras in a GTP-dependent manner, but also that single amino acid substitutions (e.g., K2150E) in the RA2 domain of PLC-ε disrupt this interaction. Similarly, both Song et al. (18) and Kelley et al. (20) illustrate that point mutations in H-Ras (e.g., N26G, Y32F, T35S, and Y40C) prevent direct interaction with the RA2 domain of PLC-ε . In contrast to RA2, the RA1 domain of PLC-ε binds H-Ras with a low affinity and in a GTP-independent manner (20) .
PLC-ε expressed ectopically in Cos-7 cells undergoes translocation from the cytosol to the plasma membrane when co-expressed with a mutant form of H-Ras deficient in GTPase activity, and to perinuclear compartments such as the Golgi apparatus when co-expressed with Rap1A (18) . Activation of the epidermal growth factor receptor on cells expressing either H-Ras or Rap1A results in membrane recruitment of GEFs, formation of H-Ras•GTP and Rap1A•GTP, and translocation of PLC-ε to either the plasma membrane in H-Ras–expressing cells or to the Golgi apparatus in Rap1A-expressing cells.
Co-expression of PLC-ε with a constitutively active mutant of H-Ras increases the rate of inositol lipid hydrolysis in Cos-7 cells (20) . This effect is attenuated by deletion of either the RA1 or RA2 domain, by mutation of RA2 (e.g., K2150E), or by mutations of H-Ras in its effector-binding domain. H-Ras•GTP activates (greater than twofold) purified PLC-ε in experiments with reconstituted liposomes (18) . Although the mechanism for H-Ras activation of PLC-ε is not clear, one interpretation of these studies is that H-Ras increases the phospholipase activity of PLC-ε by promoting the recruitment of the enzyme to the plasma membrane, that is, to the location of PtdIns(4,5)P2 substrate. The potential interplay between the RA domains and the GEF activity of the CDC25 domain (see below) has not been explored.
CDC25 Domain
Proteins that contain CDC25 domains generally act as GEFs for Ras-family GTPases, and in many instances, CDC25 domains confer specificity for particular members of the Ras family (28) . Therefore, the discovery of a CDC25 domain near the N terminus of PLC-ε is highly intriguing (17–20) . The expression of PLC-ε in TSA201 cells promotes formation of GTP•H-Ras (19) . Moreover, PLC-ε containing a single amino acid substitution (H1144L) that obliterates phospholipase activity nevertheless retains the capacity to function as a GEF, promoting formation of GTP•H-Ras. Indeed, the H1144L mutant of PLC-ε , like the wild-type enzyme and the GEF domain isolated from the wild-type enzyme, possesses the capacity to activate the mitogen-activated protein kinase pathway in a manner dependent on Ras but independent of PtdIns(4,5)P2 hydrolysis (19) .
In contrast to the results of Lopez et al. (19) , Jin and colleagues (23) report that the CDC25 domain of PLC-ε exhibits GEF activity toward Rap1 but not to other Ras-family GTPases such as H-Ras and Rap2. Purified full-length PLC-ε , or an N-terminal fragment of PLC-ε that includes the CDC25 domain (PLC-ε CDC25), increases the rate of GDP release from Rap1A, whereas a C-terminal fragment of PLC-ε (PLC-ε Δ N) has no effect. Similarly, co-expression of Rap1A with either full-length PLC-ε or PLC-ε CDC25 (but not with PLC-ε Δ N) in Cos-7 cells increases intracellular levels of Rap1A•GTP and results in activation of the downstream kinases B-Raf and extracellular signal–regulated kinase (ERK). Intracellular levels of activated Raf-1 kinase are unchanged in cells expressing PLC-ε and either H-Ras or Rap1A. Furthermore, epidermal growth factor induces the sustained translocation of PLC-ε to perinuclear compartments, such as the Golgi apparatus, in cells that express Rap1A, whereas PLC-ε lacking the CDC25 domain is only transiently translocated to these compartments (23) . These observations suggest a prominent role for the CDC25 domain of PLC-ε in amplifying Rap1-promoted signaling.
The quantification of PLC-ε –promoted inositol lipid hydrolysis and Ca2+ mobilization in response to growth factor receptor activation in Cos-7 cells is complicated by the presence of PLC-γ , which is markedly stimulated by these receptors. Therefore, to explore the molecular basis of Rap1- and H-Ras–mediated regulation of PLC-ε , Song and coworkers used platelet-derived growth factor (PDGF) to stimulate cells expressing a PDGF receptor mutant incapable of activating PLC-γ (24) . Persistent PDGF-promoted activation of ectopically expressed PLC-ε was observed, manifesting a rapid initial phase of activation mediated by Ras and a prolonged phase promoted by Rap1. The CDC25 domain, which exhibits GEF activity toward Rap1 but not H-Ras, was critical for the prolonged activation of PLC-ε . Interestingly, PDGF prevented apoptosis in BaF3 hematopoietic cells containing the mutant receptor and enabled these cells to proliferate in a PLC-ε –dependent manner. These observations are consistent with a critical role of PLC-ε for survival and growth of BaF3 cells.
Core Catalytic Domains (PH, EF, XY, C2)
Regulation of inositol lipid signaling by GPCRs had been assumed to arise entirely from the activation of PLC-β isozymes by Gα subunits of the Gq family, and in some cases, by Gβγ generated from other G protein heterotrimers. However, observations that constitutively active Gα12 (19) and Gα13 (22) , but not Gαq , enhance the phospholipase activity of PLC-ε alter this dogma. PLC-ε truncation mutants lacking the CDC25, pleckstrin homology (PH), and RA domains retain the capacity to be activated by Gα12 and Gα13 (21) . Therefore, Gα -promoted activation of PLC-ε occurs through a mechanism involving the core catalytic region (i.e., EF hands, X and Y boxes, and C2 domain) of the isozyme (Figure 3⇓). Whether the Gα12/13 -mediated activation of PLC-ε requires direct interaction of Gα and PLC-ε has not yet been determined.
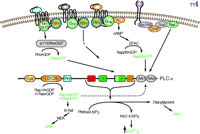
PLC-ε : A nexus for heterotrimeric and small G protein signaling pathways. Phospholipase C activity of PLC-ε is stimulated by Gα12/13, Gβγ, and activated members of Ras and Rho small GTPases to yield downstream second messengers Ins(1,4,5)P3 and diacylglycerol. In addition, ERK signaling pathways are propagated as a result of the guanine nucleotide exchange activity of the PLC-ε CDC25 domain. A positive feedback loop is schematized in which CDC25-promoted activation of H-Ras and/or Rap1A in turn activates the lipase activity of PLC-ε upon binding the C-terminal RA domains (dashed line). See text for details. (AC, adenylyl cyclase; Shc, SH2 domain–containing adaptor protein; IRS, insulin receptor substrate; Grb2, growth factor receptor–bound protein 2; Sos, son of sevenless GEF; MEK, mitogen-activated protein kinase kinase.)
RhoA, RhoB, and RhoC all markedly stimulate inositol phosphate accumulation in PLC-ε –expressing cells, whereas no effect is observed with the other members of the Rho family (e.g., Rac1, Rac2, Rac3, and Cdc42) (21) . Rho activates PLC-ε by a mechanism independent of the known small GTPase-interacting domains of the isozyme; mutant PLC-ε proteins lacking the CDC25, PH, and/or RA domains retain activation by Rho. Sequence analysis of the catalytic core of PLC-ε reveals a unique 60–70-residue insert (relative to the PLC-β , -γ , -δ , and -ζ isozymes; i.e., residues 1667–1728 of the rat PLC-ε ). A PLC-ε construct lacking this region is no longer activated by Rho or Gα12/13 but remains subject to regulation by Gβγ and H-Ras.
Gα12 and Gα13 share the capacity to activate Rho GEFs (i.e., p115RhoGEF, LARG, PDZ-RhoGEF) (29–32) , and therefore, both Gα12 and Gα13 activate Rho (33) . Although no data exist that rule out direct interaction of Gα12 and Gα13 with PLC-ε , we hypothesize that Gα12 - and Gα13 -mediated activation occurs through an intervening molecule, Rho. This speculation is supported by loss of both Rho and Gα12/13 -mediated regulation upon removal of the unique 60–70-residue sequence found in the catalytic core of PLC-ε but not other PLC isozymes (21) . Preliminary studies from our laboratory have revealed that C3 toxin, which ADP-ribosylates and inactivates Rho, prevents Gα12/13 -mediated activation of PLC-ε but has little effect on Gαq activation of PLC-β isozymes.
Both PH and EF-hand domains exist near the N terminus of PLC-ε (22) . The known interaction of Gβγ with the PH domains of other proteins (34, 35) prompted an examination of the capacity of Gβγ to activate PLC-ε . Indeed, expression of Gβ1γ2 in Cos-7 cells markedly stimulates the enzymatic activity of PLC-ε (22) ; activation is also observed with other combinations of Gβ and Gγ (e.g., Gβ1γ3 and Gβ2γ2 ). The Gβγ -promoted stimulation of PLC-ε is blocked by cotransfection with either of two Gβγ -interacting proteins, namely, Gαi1 and the C terminus of G protein receptor kinase 2. Whether the activation of PLC-ε by Gβγ occurs by a direct interaction is not known. Pharmacological inhibition of phosphoinositide 3-kinase γ does not prevent activation of PLC-ε by Gβγ . Similarly, a mutation in the RA2 domain of PLC-ε that precludes activation by Ras fails to alter the stimulatory activity of Gβ1γ2 . Although Gβγ is known to interact with certain PH domains, truncation mutants of PLC-ε lacking the PH and CDC25 domains retain Gβγ -regulated activity.
Regulation of PLC-ε through Gαs
Schmidt and coworkers (36, 37) describe a mechanism through which Gαs -coupled GPCRs potentially regulate PLC-ε . EPAC (for e xchange p rotein a ctivated by c AMP), a Rap-specific GEF, potentiates the inositol trisphosphate and Ca2+ responses to GPCR stimulation by catecholamines and prostaglandins. Therefore, these investigators examined the roles of Rap GTPases and PLC-ε in Gαs -coupled GPCR signaling. Dominant negative forms of Rap1A, 1B, 2A, and 2B, as well as a dominant negative form of H-Ras, were expressed in HEK-293 cells with the goal of blocking Ins(1,4,5)P3 accumulation in response to β -adrenergic receptor stimulation. The dominant negative form of Rap2B, but not the other dominant negative constructs, inhibited the isoproterenol- and forskolin-promoted formation of Ins(1,4,5)P3 . GTP-dependent binding of Rap2B to PLC-ε RA2 has been demonstrated (24) , and thus, cyclic AMP–promoted activation of PLC-ε conceivably occurs through activation of Rap2B and interaction with the RA2 domain (Figure 3⇑).
Conclusion
The observations made by Cantley and coworkers (38) and Wakelam et al. (39) nearly twenty years ago suggested important roles for Ras in the regulation of PtdIns(4,5)P2 signaling. Recent revelations in the study of the novel PLC isozyme, PLC-ε , have resurrected this notion and opened a broad set of questions for researchers in the increasingly integrated fields of heterotrimeric and small G protein signaling. The dogma that signaling through heterotrimeric G proteins and through members of the Ras superfamily of small GTPases occurs in a linear and unidirectional fashion thus continues to be refined by the realization that significant crosstalk occurs between these complex signaling pathways.
PLC-ε exhibits unique regulatory potential as both an initiator and recipient of activated G protein signals. A positive feedback loop also may operate through the isozyme in which H-Ras and/or Rap become GTP-bound through interaction with the CDC25 domain and subsequently activate the phospholipase activity of PLC-ε via interaction with the RA2 domain, enabling long-lasting signal maintenance. Given these unique signaling attributes and wide tissue distribution of PLC-ε –encoding mRNA, we assume PLC-ε subserves critical roles in mammalian physiology. PLC-ε is well placed to function in the regulation of cell morphology, development, and proliferation, and in this way most assuredly merits attention as a potential therapeutic target.
- © American Society for Pharmacology and Experimental Theraputics 2003
References

Ken Harden, Ph.D. (left), is Kenan Professor of Pharmacology at the University of North Carolina School of Medicine. David Bourdon, Ph.D. (middle), and Michele Wing, Ph.D. (right), are postdoctoral fellows in the Department of Pharmacology at the University of North Carolina.

