|
|
Variantes des gènes de l’adiponectine (ADIPOQ) et du récepteur 1 de l’adiponectine (ADIPOR1) et risque de cancer colorectal
Virginia G. Kaklamani, MD, DSc;
Kari B. Wisinski, MD;
Maureen Sadim, BS;
Albert Do, BS;
Kenneth Offit, MD, MPH;
John A. Baron, MD;
Habibul Ahsan, MD, MPH;
Christos Mantzoros, MD, MPH;
Boris Pasche, MD, PhD
JAMA. 2008;300(13):1523-1531
RÉSUMÉ
| |
Contexte Les preuves épidémiologiques du moment suggèrent qu'il existe un lien entre l'obésité, l'hyperinsulinémie et le risque de cancer colorectal. L'adiponectine est une hormone sécrétée par les tissus adipeux, et les taux de sérum sont inversement proportionnels à l'obésité et à l'hyperinsulinémie. Alors qu'il y a des preuves de lien entre les taux de circulation d'adiponectine et le risque de cancer colorectal, aucun lien entre les gènes de l'itinéraire de l'adiponectine et le risque de cancer colorectal n'a été noté.
Objectif Déterminer le lien qui existe entre 10 polymorphismes à noyau unique marqueurs d'haplotype de l'adiponectine (ADIPOQ) et du récepteur d'adiponectine 1 (ADIPOR1) et le risque de cancer colorectal.
Modèle, contexte et patients Deux études de cas ont été utilisées, elles comprenaient des patients ayant ou non un diagnostique de cancer colorectal recruté entre 2000 et 2007. L'étude de cas 1 comprenait 441 patients diagnostiqués de cancer colorectal et 658 cas négatifs; les deux groupes étaient formés de personnes Juives Ashkénazes vivant à New York City, New York. L'étude de cas 2 comprenait 199 patients diagnostiqués de cancer colorectal et 199 patients cas négatifs provenant de Chicago, Illinois; les individus ont été mis par paires d'âge, de sexe et de race.
Principales mesures de résultat Les fréquences d'ADIPOQ et d'ADIPOR1 parmi les cas positifs et négatifs.
Résultats Dans l'étude 1, et après ajustement de l'âge, du sexe et de SNP du même gène, 3 SNP d'ADIPOQ et 1 SNP d'ADIPOR1 ont été associés à un risque de cancer colorectal: rs266729 (rapport de cotes ajusté [AOR], 0.72; intervalle de confiance [IC,], 0.55-0.95) et rs822396 (AOR, 0.37; 95% IC, 0.14-1.00) ont été associés à un risque moindre alors que rs822395 (AOR, 1.76; 95% IC, 1.09-2.84) et rs1342387 (AOR, 1.79; 95% IC, 1.18-2.72) ont été associés à un risque accru. Dans l'étude 2, après ajustement sur l'âge, le sexe, la race et les SNP du même gène, le SNP d'ADIPOQ rs266729 a été associé à un risque de cancer colorectal moindre ou de magnitude similaire à celui de l'étude 1 (AOR, 0.52; 95% IC, 0.34-0.78). L'analyse des deux études combinées montre un lien entre le rs266729 et un risque moindre de cancer colorectal (AOR, 0.73; 95% IC, 0.53-0.99).
Conclusions Le SNP rs266720, qui marque la 5' flanking région du gène ADIPOQ, est associé à un risque moindre de cancer colorectal.
Plusieurs études épidémiologiques ont montré qu'il existait un lien entre l'adiposité et le risque de cancer colorectal incident 1-5. Des études d'observation ont également montré que des marqueurs de résistance à l'insuline, tels que des taux de circulation de C-peptide ou d'IGFBP1 (insulin-growth factor-binding protein 1) élevés, sont directement associés à un risque de cancer colorectal 6-9.
L'adiponectine est une protéine sécrétée par le tissu adipeux et il a été prouvé qu'elle est un sensibilisateur d'insuline endogène. A la différence d'autres adipokines telles que la leptine, les taux de circulation d'adiponectine sont plus bas chez les personnes obèses et ceux touchées de diabète 10. Etant donné le lien entre l'adiponectine et la résistance à l'insuline et à l'IGF ainsi que le lien entre la résistance à l'insuline et à l'IGF et le risque de cancer colorectal, nous avons posé comme hypothèse que l'itinéraire de l'adiponectine pourrait influer directement sur le risque de cancer colorectal.
Plusieurs types de preuves soutiennent cette hypothèse. L'adiponectine et ses récepteurs sont rapidement dirigés vers le tissu colonique 11,12. Il semble que l'adiponectine agisse dans les lésions coloniques prénéoplastiques afin de réguler la croissance cellulaire en activant, alternant ou interagissant avec plusieurs itinéraires y compris ceux de la leptine et de la NF-kB 13. De plus, nous avons précédemment montré que les taux d'adiponectine sont inversement liés au risque de cancer colorectal 14. Dans une étude prospective de contrôle de cas isolée, les hommes ayant des taux d'adiponectine bas couraient un risque de cancer colorectal plus grand que ceux ayant des taux bas. Comparés avec les hommes du quintile le plus bas, les hommes du quintile d'adiponectine le plus haut avaient 68% de moins de chances de développer un cancer colorectal (risque relatif, 0.42; 95% Intervalle de Confiance [IC, Confidence Interval], 0.23-0.78) 14. En outre, les taux d'expression d'ADIPOR1 et d'ADIPOR2, les deux récepteurs d'adiponectine, sont supérieurs dans les cas de carcinomes colorectaux en comparaison avec des épithéliums coloniques normaux 12.
Il a été démontré que plusieurs polymorphismes d'adiponectine influencent les taux d'adiponectine et le polymorphisme de gènes car le ligand (ADIPOQ, ID du gène 9370, GeneBank ID NM_004797) et son récepteur de type 1 (ADIPOR1, ID du gène 51094, GeneBank ID NM_015999) ont été associés à un risque de résistance à l'insuline, à des maladies cardiovasculaires et au diabète sucré 15-22. Les rôles de résistance à l'insuline des gènes récepteur d'adiponectine 1 (ADIPOR1) et récepteur d'adiponectine 2 (ADIPOR2) ont été évalués par plusieurs associations d’études génétiques. Crimmins et Martin 22 ont résumé ces découvertes et ont conclu qu'un seul polymorphisme d'ADIPOR1 à nucléotide unique (SNP, single-nucleotide polymorphism) (rs134287) était associé de façon significative au risque de résistance à l'insuline. Le lien entre ces polymorphismes et le risque de cancer colorectal n'a cependant pas été étudié. Afin d'examiner le lien entre deux gènes clés de l'itinéraire de l'adiponectine et du risque de cancer colorectal, nous avons généotypé 10 SNP marqueurs d'haplotype des gènes ADIPOQ et ADIPOR1 au cours de deux études de cas indépendantes.
METHODES
Participants à l'étude
La première étude a été conduite dans le but d'évaluer les liens potentiels entre les polymorphismes d'ADIPOQ et de ADIPOR1 et les risques de cancer colorectal. La seconde étude de cas a été utilisée à des fins de réplication des découvertes de la première.
Étude de cas 1
En tant que partie intégrante des protocoles approuvés par le comité institutionnel de révision (IRB, institutionnal review board), nous avons obtenu des échantillons de sang provenant de 441 patients hospitalisés au Memorial Sloan-Kettering Cancer Center (New York City, New York) diagnostiqués d'un cancer colorectal. Une accumulation de cas consécutifs est apparue dans la période du 1er Avril 2000 au 31 Décembre 2000, et du 1er Octobre 2005 au 28 Février 2006. Tous les cas de cancer colorectal ont été histologiquement confirmés au Memorial Sloan-Kettering Cancer Center. Les informations au sujet du sexe, de l'âge, de l'âge au moment du diagnostic du cancer et de la race ont été enregistrées. Un échantillon de 658 volontaires en bonne santé venant de New York City, New York a été recruté entre le premier Janvier 2003 et le 31 Décembre 2004 en tant que sous-groupe du New York Cancer Project, une grande cohorte d'étude ayant été précédemment décrite en détail 23. Le taux de réponse était de 89%. Aucun cas négatif n'avait de passé cancéreux au moment du don de sang, ce qui fut confirmé par un questionnaire complété par chaque volontaire en bonne santé. Tous les participants ont signé une feuille de consentement informée approuvée par l'IRB. Tous les cas positifs et négatifs étaient blancs et Juifs Ashkénazes.
Étude de cas 2
La seconde étude de cas comprenait 199 paires assorties de cas positifs et négatifs de cancer colorectal provenant de la Northwestern University à Chicago, Illinois. Les cas positifs et négatifs étaient mis en paires d'âge, de race et de sexe. La race a été retenue comme étant une variable car le risque de cancer varie selon le groupe ethnique. Des cas positifs consécutifs ayant eu un diagnostique d'adénocarcinome colorectal confirmé par biopsie ont été recrutés par des cliniques médicales et de chirurgie oncologique affiliées à la Northwestern Medical Faculty Foundation and US Oncology en deux phases (1er Juin 2006 au 31 Août 2006 et 15 Janvier 2007 au 31 Mai 2007). Les taux de réponses étaient respectivement de 92% et 86%, et on a obtenu un échantillon sanguin de chaque patient recruté. Tous les cas positifs ont signé une feuille de consentement informée sur les études génétiques et le protocole a été approuvé par l'IRB de la Northwestern University. Les cas négatifs ont été choisis parmi un total de 5578 patients sans diagnostique de cancer au moment de l'inscription qui ont été recrutés entre le 31 Octobre 2002 et le 31 Décembre 2007. Les cas négatifs ont été mis par paires d'âge à cinq ans près, par sexe et par race. Les cas négatifs provenaient du Nugene Project, un dépôt de biospécimen en convention avec le Health Insurance Portability and Accountability Act et approuvé par l'IRB, et qui comprend des données de questionnaires ainsi que des information médicales longitudinales provenant des patients d'hôpitaux et de cliniques affiliées à la Northwestern University.
Les participants potentiels ont été approchés par un conseiller génétique ou par un médecin. Le taux de réponse global a été 25%. Les deux raisons principales de refus de participer étaient un manque de temps ou des soucis de confidentialité. Le dépôt représente une population clinique diverse avec des échantillons de logement et des patients souffrant de pathologies les plus communes telles que le diabète, le cancer et des maladies auto immunitaires et cardiovasculaires. Les participants ont signé une feuille de consentement informée approuvée par l'IRB afin de rendre possibles la distribution et l'utilisation d'échantillons à identité enlevée et de données provenant d'un large panel de recherche. La possibilité de mettre à jour régulièrement les états de santé des participants permet à Nugene d'offrir aux chercheurs les échantillons les plus pertinents à leur recherche. Les cas négatifs ont été sélectionnés parmi les participants de Nugene n'ayant fait état d'aucun passé de cancer. Les patients présentant des codes d'International Classification of Diseases, Ninth Revision indiquant un diagnostic de cancer ont été exclus.
Isolement d'ADN
L'ADN de lymphocytes sanguins entiers a été extrait en se servant du QIAamp DNA Blood Mini Kit (Qiagen, Hilden, Germany) et a été stocké à -20°C jusqu'à son utilisation pour le génotypage. Tous les échantillons d'ADN ont été soumis à des amplifications de génome entier en se servant du Illustra Genomiphi V2 DNA Amplification Kit (GE Healthcare, Waukesha, Wisconsin; catalog No 25660032). Les échantillons étaient stockés à -20°C.
Sélection des SNP
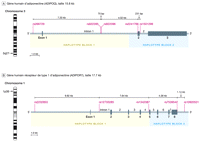
Nous avons sélectionné les SNP en capturant les variations au sein de blocs majeurs de chaque gène. Nous avons cependant préféré choisir les SNP pertinents fonctionnellement parmi ces SNP marqueurs et avons par conséquent généotypé 10 SNP qui affectent les taux d'adiponectine ou qui sont liés à un risque de résistance à l'insuline, de maladies cardiovasculaires ou de diabète 15-22. De plus, nous n'avons sélectionné que les SNP présentant une fréquence allélique de 10% parmi les blancs. Le gène d'adiponectine a plus de 10 SNP marqueurs 16,21 et deux blocs de déséquilibre de liens avec une attache de bloc comprise entre -2049 et -450 18. Nous avons choisi de généotyper rs266729 (5' flanking region), rs822395 (intron 1) et rs822396 (intron 1) pour marquer le bloc 1 et rs1501299 (intron 2) et rs2241766 (exon 2) pour marquer les bloc 2 puisque ces 5 SNP sont les plus communes et ont été étudiés le plus exhaustivement par d'autres de par leur fonctionnalité et leur lien avec des maladies telles que le diabète sucré 15-18 (Figure). L'ADIPOR1 possède plus de 28 SNP dans deux blocs de déséquilibre de liens 20. L'un des blocs s'étend de la 5' flanking region à l'intron 4 et l'autre est situé à l'extrémité 3' du gène. En se basant sur cette structure, nous avons sélectionné 5 SNP communs pour le génotypage. Pour le bloc 1, nous avons sélectionné les marqueurs SNP suivants: rs2232853 (5' flanking region), rs12733285 (intron 1), et rs1342387 (intron 4). Pour le bloc 2, nous avons sélectionné rs7539542 (exon 8) et rs10920531 (3' flanking region). Ces SNP ont été sélectionnés car ils marquent les deux blocs de déséquilibre de liens.
|
|
|
Figure. Haplotypes du gène d'adiponectine et polymorphismes marqueurs à noyau unique
A- Le gène d'adiponectine (ADIPOQ) se rend au 3q27 et contient 2 blocs d'haplotype. Quatre SNP introniques et un SNP exonique ont été sélectionnés pour génotyper ces deux blocs. B- Le gène récepteur de type 1 d'adiponectine (ADIPOR1) se rend au 1p36 et contient 2 blocs d'haplotype. Quatre SNP introniques et un SNP exonique ont été sélectionnés pour génotyper ces deux blocs. La désignation des blocs d'haplotypes dans la Figure est approximative.
|
|
|
Nous nous sommes servis des sondes VIC et FAM. Les amorces des différents SNP pour ADIPOQ et ADIPOR1 sont montrées dans l'Encadré 1.
| Encadré 1. Polymorphismes à noyau unique pour ADIPOQ et ADIPOR1 Amorces d'ADIPOQ
rs266729: TTGCAAGAACCGGCTCAGATCCTGC[C/G]CTTCAAAAACAAAACATGAGCGTGC
rs822395: TGATCGCACCTATTAGTGGAGAAAT[A/C]TGTCCATAATACTGAAGTTTGGGGA
rs822396: GTAGGAGAAAGAGATCTTTATTTTT[A/G]ACAAAGGGGGAGGAGCCAGAAAACT
rs2241766: TTCTACTGCTATTAGCTCTGCCCGG[G/T]CATGACCAGGAAACCACGACTCAAG
rs1501299: CTACACTGATATAAACTATATGAAG[G/T]CATTCATTATTAACTAAGGCCTAGA
Amorces d'ADIPOR1
rs2232853: CAAGTGGTAGCAGCAGCTGGGAAT[A/G]GGTATACTCAGCCTGCCTCAAGCTG
rs12733285: TCATGCTATGCTCAACCCACAAGCA[C/T]AGTTGAAAGCAACCGGCAATCTAGT
rs1342387: AAAAAAGGGAATGTGTACACTTTGA[C/T]GGTTGATGTTTTTGAATCAGAGAGC
rs7539542: CATGTGAAATCTTTGAATGCCAAGT[C/G]TCTTCTGTACTTTCTTTTATTAACA
rs10920531: AAACTTGACTCTTGACATGAACCCA[A/C]CTTTAACTCAAAAAGACTGCCCTTA
|
|
Génotypage
Le génotypage des 10 SNP a été effectué par discrimination allélique de SNP Taqman avec un ABI 7900HT (Applied Biosystems, Forest City, California). Les résultats ont été confirmés par la version 2.3 du logiciel SDS (Applied Biosystems). Tous les résultats étaient automatiquement appelés (c'est à dire que l'appareil montre automatiquement les génotypes avec une certitude de 95%). Un total de 5% des échantillons a été génotypé en double et étaient concordants à 100%.
Analyse statistique
Nous nous sommes servis de Tests X² pour comparer la fréquence et les distributions démographiques des génotypes parmi les cas positifs et négatifs.
Nous nous sommes servis de régressions logistiques inconditionnelles à variantes uniques ou multiples afin d'évaluer les Odds Ratio (OR, rapports de cotes) bruts et ajustés. Des modèles ajustés de régression logistiques à variante multiple ont été ajustés à chaque SNP en fonction de l'âge, du sexe et d'autres SNP du même gène. Plus spécifiquement pour l'étude 1, des OR bruts ont été ajustés en fonction de l'âge (continu), du sexe (binaire), et des SNP du même gène (catégorique). Pour l'étude de cas 2, les OR bruts ont été ajustés pour correspondre aux facteurs d'âge (continu), de sexe (binaire) et de race (catégorique), d'autres OR ont été en plus ajustés aux SNP du même gène (catégorique). Pour l'analyse combinée, les OR bruts n’étaient pas ajustés mais d'autres OR l'étaient à l'âge (continu), au sexe (binaire), à la race (catégorique), et à d'autres SNP du même gène (catégorique).
Les modèles additifs et dominants ont été utilisés pour l'analyse. Dans le modèle additif, chaque SNP est modelé catégoriquement et séparé en 3 catégories, un pour chaque génotype avec un génotype désigné comme étant le groupe de référence. Pour le modèle dominant, chaque SNP est modelé comme une variable dichotomique, un génotype est désigné comme étant le groupe de référence et les deux autres sont combinés dans une catégorie.
Toutes les comparaisons étaient à deux facettes et ont été effectuées avec une valeur de P de .05. En se servant d'une prévalence de .48 (prévalence de GG/CG pour SNP rs266729 dans les cas négatifs), nous avons calculé que la puissance nécessaire pour détecter un OR de 2 était comme suit: étude combinés, puissance=0.99; étude assortie (cas positifs et négatifs), puissance=0.93; et étude non assortie (étude de cas 1), puissance=0.99. Nous n'avons effectué aucun ajustement aux comparaisons multiples dans l'analyse. Toutes les valeurs de P peuvent être multipliées par 10 afin de donner des valeurs de P ajustées au Bonferroni et les niveaux de signification à deux facettes peuvent être remplacées par .005. L'analyse statistique a été faite avec le logiciel de statistique SAS version 9.1 (SAS Institute, Inc., Cary, North Carolina).
RESULTATS
Démographie
Les cas positifs et négatifs de l'étude 1 étaient tous des personnes blanches résidant à New York City, New York (Tableau 1). L'âge moyen des cas positifs était de 64 and (étendue interquartile [Interquartile Range, IQR], 55.0-71.0 ans) et l'âge moyen des cas négatifs était de 51 ans (IQR, 43.0-58.6 ans). Il y avait plus d'hommes parmi les cas positifs que négatifs. Les individus de l'étude de cas 2 étaient mis par paires de race, d'âge (à cinq ans près), de situation géographique et de sexe (Tableau 1). L'âge moyen des cas positifs était de 60 ans (IQR, 51.0-69.0 ans) et l'âge moyen des cas négatifs était de 58 ans (IQR, 49.0-67.0 ans).
|
|
|
|
Tableau 1. Démographie des deux études de cas
|
|
|
SNP ADIPOQ et risque de cancer colorectal
Les Tableaux 2 et 3 présentent les résultats respectivement sous les modèles additifs et dominants. L'analyse sous le modèle dominant a montré que 3 SNP marqueurs d'haplotype étaient associés à des risques de cancer colorectal dans l'étude de cas 1 (Tableau 2). Les génotypes rs266729 GG/CG ont été associés à un risque de cancer colorectal moindre (OR, 0.72; IC 95%, 0.55-0.95). Les génotypes rs822395 AA/AC ont été associés à une augmentation du risque de cancer colorectal (OR, 1.76; IC 95%, 1.09-2.84) et les génotypes rs822396 AA/AG ont révélé un risque de cancer colorectal moindre (OR, 0.37; IC 95%, 0.14-1.00). Dans l'étude de validation (étude de cas 2), seul le polymorphisme rs266729 a été associé de façon significative à un risque (risque relatif pour les génotypes GG/CG, 0.52; IC 95%, 0.34-0.78). Les deux analyses combinées sous le modèle dominant sont présentées dans le Tableau 4. Les fréquences allèles du rs266729 parmi les cas négatifs sont comparables dans les deux études de cas: GG, 6% et 7%; CC, 41% et 51%; et CG, 52% et 41%. Après ajustement à l'âge, au sexe, à la race et aux SNP des mêmes gènes, les rs266729 a été associé à un risque de cancer colorectal réduit (OR, 0.73; IC 95%, 0.53-0.99). Après ajustement à variante multiple, il subsiste un lien négligeable entre le rs822395 et le risque de cancer colorectal (OR, 0.79; IC 95%, 0.61-1.02; P=.07). Rs266729 et rs822395 appartiennent tous les deux au même bloc d'haplotype (Figure). Chaque SNP lié à un risque de cancer colorectal est en adéquation avec l'équilibre de Hardy-Weinberg en se qui concerne les cas négatifs.
|
|
|
|
Tableau 2. Modèle dominant des rapports de cotes bruts et ajustés (OR) par des génotypes de polymorphismes à noyau unique d'ADIPOQ et de ADIPOR1
|
|
|
|
|
|
|
Tableau 3. Modèle dominant des rapports de cotes bruts et ajustés (OR) de cancer colorectal par des génotypes de polymorphismes à noyau unique d'ADIPOQ et de ADIPOR1
|
|
|
|
|
|
|
Tableau 4. Analyse combinée sous le modèle dominant
|
|
|
SNP ADIPOR1 et risque de cancer colorectal
5 SNP marqueurs d'ADIPOR1 ont été génotypés. Dans l'étude de cas 1, les génotypes rs1342387 CC/TC étaient liés à un risque accru de cancer colorectal (OR, 1.79; IC 95%, 1.18-2.72). Ce lien n'a pas été confirmé par l'étude de validation (OR, 1.06; IC 95%, 0.68-1.66). L'analyse combinée des deux études sous le modèle dominant ne montre aucune association entre les SNP ADIPOR1 et le risque de cancer colorectal (Tableau 4). Chaque SNP associé à un risque de cancer colorectal est en adéquation avec l'équilibre Hardy-Weinberg en ce qui concerne les cas négatifs.
COMMENTAIRE
Dans cette étude de cas clinique, nous avons découvert l'existence d'un lien entre un SNP du gène ADIPOQ (rs266729) et le risque de cancer colorectal dans deux études de cas distinctes ainsi que dans une analyse combinée des deux études après un ajustement en fonction de l'âge, du sexe, et d'autres SNP.
Il existe des preuves considérables allant dans la direction de l'importance du signalement d'adiponectine dans la carcinogenèse colorectale. Les tissus coloniques transitent l'ADIPOR1 et l'ADIPOR2 12. Deux études récentes ont démontré d'une association entre les taux de sérum d'adiponectine et le risque de cancer colorectal 12,14, avec une réduction du risque de 47% dans le quintile le plus élevé en comparaison avec le quintile le plus bas 14. D'autres recherches n'ont cependant pas conclu par cette association 24.
Le lien entre l'adiponectine et le cancer colorectal pourrait être l'itinéraire IGF. L'hyperinsulinémie réduit les taux de circulation d'IGFBP1, ce qui pourrait provoquer une augmentation des taux d'IGF1 non lié. Des taux élevés d'IGF1 en circulation, ce qui accroît la prolifération cellullaire et inhibe l'apoptose 25,26, ont été associés à un risque plus important de plusieurs cancers communs, y compris la cancer colorectal 6,27,28. Nous avons également récemment exposé qu'un taux d'insuline élevé (en reflet d'une augmentation de C-Peptide) ou qu'un taux de IGF1 biodisponible élevé (évalué par le ration IGF1/IGFBP3) prédisait indépendamment d'un risque accru de cancer colorectal; des taux élevés des deux n'étaient pas associés à un risque accru 9.
Dans les deux études de cas incluant un total de 1497 personnes, nous avons observé une association constante entre le rs266729 et le risque de cancer colorectal. Pour être plus précis, les personnes ayant un génotype GG/GC ont 27% de moins de chances de contracter un cancer colorectal que ceux du génotype CC. Il a été démontré que l'allèle G est associé à des taux d'adiponectine moindres 19. Le SNP marqueur d'haplotype rs266729 est situé sur la 5' flanking region du gène à une position de -11365 dans la région du promoteur. Les polymorphismes de la région du promoteur ont été associés aux taux d'adiponectine et au risque de diabète 21. De plus, rs266729 a été associé à un risque de pontage coronarien et d'angioplastie coronaire transluminale percutanée 29. D'autres polymorphismes de la même région, y compris rs2117985 (-18003), rs822387 (-14811) et rs860291 (-12891) ont également été associés de façon significative aux taux d'adiponectine 17. Des personnes avec des génotypes rs17300539 (-11391) AA/AG ont en outre des taux d'adiponectine supérieurs de 64% (P=.01) 18. Dans l'étude que nous exposons, nous n'avons choisi d'étudier qu'un seul polymorphisme de la 5' flanking region. Nos données ainsi que celles obtenues d'autres chercheurs mettent l'accent sur l'importance de cette région au sujet de la fonction du gène ainsi que du fait qu'elle est une région encline à causer des maladies du gène d'adiponectine. Il est intéressant de constater que l'analyse combinée des deux études présentes dans cet article ont démontré d'un lien contigu entre rs822395 et le risque de cancer colorectal, ce qui suggère que plusieurs SNP de cette région pourraient être liés à des risques de cancer colorectal.
Nous avons récemment évalué 10 SNP de l'itinéraire de l'adiponectine en relation avec le risque de cancer du sein 30. Nos découvertes ont montré qu'une région différente du gène d'adiponectine était un modificateur important du risque de cancer du sein. Plus spécifiquement, nous avons découvert que +45 T–>G 9rs2241766) et que +276 G–>T (rs1501299) sont associés au risque de cancer du sein. Ces deux polymorphismes appartiennent au bloc 2; rs2241766 est situé dans l'exon 2 alors que rs1501299 est dans l'intron 2. Nous avons découvert qu'un polymorphisme d'ADIPOR1 (+10225 C–>G [rs7539542]) est associé de façon significative au risque de cancer du sein. Les SNP marqueurs d'haplotype utilisés dans cette étude représentent des régions génomiques putativement associées au risque de cancer du sein mais les SNP et/ou mutations fonctionnellement pertinentes sont sans doute différents de ces SNP marqueurs d'haplotype. Il sera nécessaire de produire des études supplémentaires dans le futur afin de savoir si les SNP fonctionnels associés au risque de cancer du sein diffèrent de ceux associés au cancer colorectal.
Cette étude comporte a plusieurs points forts. L'étude de cas 1 incluait un large nombre de cas positifs et négatifs (N=1099) de la même race et provenant de lieux identiques. Tous les cas inclus dans l'étude de cas 2 ont été recrutés dans la même institution et ont été mis par paires d'âge, de sexe et de race. L'une des quatre combinaisons génotypiques associés au cancer colorectal dans l'étude 1 a été confirmée dans l'étude 2 malgré les différences d'âge, de race et malgré la proportion d'hommes et de femmes dans les deux études. La magnitude du lien entre rs266729 et le cancer colorectal était comparable dans les deux études. De plus, une analyse combinée des deux études a montré que le lien pertinent entre rs266729 et le cancer colorectal persistait après les ajustements d'âge, de sexe, de race et de SNP sur le même gène. En se servant d'une prévalence de 0.48 (prévalence de GG/CG du SNP rs266729 pour les cas négatifs), notre étude avait une portée suffisante pour détecter un OR de 2 (étude combinée: portée=0.99; étude assortie: portée=0.93; étude non assortie, portée=0.99).
Alors que rs226729 était associé à un risque de cancer colorectal dans les deux études, ce SNP ne représente qu'une région génomique putativement associées à un risque. Il est probable que les SNP et mutations fonctionnellement significatives soient différents. Il faudra d'autres études incluant le reséquençage de cette région génomique et des analyses de SNP dense pour identifier ces SNP ou mutations et déterminer leur impact sur les taux d'expression d'ADIPOQ.
Notre étude comporte également plusieurs limites. Le modèle épidémiologique est relativement informel, avec des populations de cas positifs et négatifs différentes. Les cas positifs et négatifs de l'étude 1 ont différé de façon significative au niveau du sexe et de l'âge car l'âge moyen des cas positifs était légèrement plus élevé que pour les cas négatifs. Il se peut que les différences d'âge parmi les cas négatifs et les positifs aient affecté les fréquences allèles observées. De plus, des informations non correctes en rapport avec toute classification par variable seraient rendues caduques et non-différentielles, ce qui à son tour mènerait à un biais conservateur dans les mesures d'association. Une limite commune aux deux études est l'absence d'information sur le passé de cancer colorectal dans la famille ainsi que tout autre risque de cancer colorectal. Une autre limite réside dans le fait que les deux études diffèrent dans les populations qu'elles ont étudié. Tous les cas positifs et négatifs étaient de race blanche et Juifs Ashkénazes dans l'étude 1, alors que l'étude 2 incluait plusieurs groupes ethniques. Les SNP en sus associés à un risque de cancer colorectal dans l'étude 1 pourraient être propres aux Juifs Ashkénazes, une population à risque de cancer colorectal accru. APCI1307K, un gène associé à un risque accru de cancer colorectal chez les Juifs Ashkénazes, se rend au 5q21-22. Si un ou plus des 3 SNP associés au cancer colorectal dans l'étude 1 venait à être validé au sein d'une autre population de Juifs Ashkénazes, il serait alors intéressant d'étudier les interactions gène-gène entre ces SNP et APCI1307K. Nous n'avons en outre pas formellement ajusté nos résultats à la multitude de comparaisons faites avec les 10 SNP. Pourtant, cet ajustement n'aurait pas matériellement changé les conclusions de notre étude.
Cette étude est à notre connaissance la seule faisant état d'un lien entre des polymorphismes et l'itinéraire de l'adiponectine, et le risque de cancer colorectal. Ce n'était pas le but de notre étude d'apporter une évaluation exhaustive de tous les gènes présents sur l'itinéraire de l'adiponectine. Nous découvertes suggèrent cependant que l'ADIPOQ rs266729 est lié à un risque de cancer colorectal. Il est important de noter qu'elles suggèrent que la 5' région du gène ADIPOQ porte des SNP et mutations susceptibles de modifier le risque de cancer colorectal. Si ces résultats peuvent être confirmés par d'autres études, l'axe de l'adiponectine pourrait émerger comme étant un modificateur important du risque de cancer colorectal. Il faudra que dans le futur d'autres études se penchent sur l'impact potentiel de l'adiponectine et de ses SNP sur le pronostique de cancer colorectal et qu'elles soient même éventuellement incorporées aux modèles de risque génétiques de cette maladie.
Informations sur les auteurs
Correspondance : Boris Pasche, MD, PhD, Division of Hematology/Oncology and Comprehensive Cancer Center, University of Alabama, 1802 Sixth Ave S, NP 2666, Birmingham, AL 35294 (Boris.Pasche{at}ccc.uab.edu).
Contributions des auteurs : Le Dr Pasche avait un accès illimité a toutes les données de l'étude et endosse toute responsabilité au sujet de l'intégrité des données et de la précision des analyses de données.
Concept et modelage de l'étude: Kaklamani, Sadim, Ahsan, Mantzoros, Pashe.
Acquisition des données: Kaklamani, Sadim, Gulden, Offit
Analyse et interprétation des données: Wisinski, DO, Barin, Ahsan, Mantzoros, Pasche.
Elaboration du manuscrit: Kaklamani, Do, Ahsan, Mantzoros, Pasche.
Relecture critique du manuscrit pour les contenus intellectuels importants: Kaklamani, Wisinski, Sadim, Gulden, Offit, Baron, Ahsan, Mantzoros, Pasche.
Analyse statistique: Kaklamani, Sadim, Do, Mantzoros.
Financements obtenus: Ahsan, Pasche.
Aide administrative, technique et matérielle: Offit, Baron.
Supervision de l'étude: Kaklamani, Ahsan, Mantzoros, Pasche.
Dévoilement de financements: Aucun.
Financement/Aide: Cette étude a été soutenue par la Walter S. Mander Foundation (Chicago, Illinois); par les bourses CA112520, CA108741, DK081913, DK79929 et DK58785 du National Institutes of Health; par la bourse Jeannick M. Littlefield for Metastatic Colon Cancer Research du American Institute for Cancer Research; par une bourse non dévoilée du Beth Israel Deaconess Medical Center; par le prix Lynn Sage Foundation and the American Society of Clinical Oncology Career Developpement; et par la Niehaus Clinical Cancer Genetics Initiative.
Rôle des sponsors: les organisations de financement n'ont eu aucun rôle dans la construction de modèle ou dans la conduite de l'étude, ni dans la récupération, prise en charge, analyse et interprétation des données; et non plus dans la préparation, relecture ou approbation du manuscrit.
Dégagement de responsabilité: Le Dr Pasche est un Contributing Editor du JAMA mais n'a pas une fois été impliqué dans les décisions d'évaluation ou dans les décisions éditoriales en ce qui concerne la publication de cet article.
Affiliations d’Auteurs : Cancer Genetics Program, Division of Hematology and Oncology, Department of Medicine and Robert H. Lurie Comprehensive Cancer Center, Feinberg School of Medicine, Northwestern University, Chicago, Illinois (Drs Kaklamani, Wisinski, and Pasche, and Mss Sadim and Gulden); Division of Endocrinology and Metabolism, Department of Medicine, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, Massachusetts (Mr Do and Dr Mantzoros); Clinical Genetics Service, Department of Medicine, Memorial Sloan-Kettering Cancer Center, New York, New York (Dr Offit); Departments of Medicine and Community and Community and Family Medicine, Dartmouth Medical School, Hanover, New Hampshire (Dr Baron); and Department of Health Studies, Medicine, and Human Genetics, University of Chicago, Chicago, Illinois (Dr Ahsan). Dr Pasche is now with the Division of Hematology/Oncology and Comprehensive Cancer Center, University of Alabama, Birmingham.
BIBLIOGRAPHIE
| |
1. Lee IM, Paffenbarger RS Jr. Quetelet’s index and risk of colon cancer in college alumni. J Natl Cancer Inst. 1992;84(17):1326-1331.
FREE FULL TEXT
2. Calle EE, Rodriguez C, Walker-Thurmond K, Thun MJ. Overweight, obesity, and mortality from cancer in a prospectively studied cohort of US adults. N Engl J Med. 2003;348(17):1625-1638.
PUBMED
3. MacInnis RJ, English DR, Hopper JL, Haydon AM, Gertig DM, Giles GG. Body size and composition and colon cancer risk in men. Cancer Epidemiol Biomarkers Prev. 2004;13(4):553-559.
FREE FULL TEXT
4. Pischon T, Lahmann PH, Boeing H, et al.. Body size and risk of colon and rectal cancer in the European Prospective Investigation Into Cancer and Nutrition (EPIC). J Natl Cancer Inst. 2006;98(13):920-931.
FREE FULL TEXT
5. Larsson SC, Wolk A. Obesity and colon and rectal cancer risk: ameta-analysis of prospective studies. Am J Clin Nutr. 2007;86(3):556-565.
FREE FULL TEXT
6. Giovannucci E, Pollak MN, Platz EA, et al.. A prospective study of plasma insulin-like growth factor-1 and binding protein-3 and risk of colorectal neoplasia in women. Cancer Epidemiol Biomarkers Prev. 2000;9(4):345-349.
FREE FULL TEXT
7. Ma J, Giovannucci E, Pollak M, et al.. A prospective study of plasma C-peptide and colorectal cancer risk in men. J Natl Cancer Inst. 2004;96(7):546-553.
FREE FULL TEXT
8. Sandhu MS, Dunger DB, Giovannucci EL. Insulin, insulin-like growth factor-I (IGF-I), IGF binding proteins, their biologic interactions, and colorectal cancer. J Natl Cancer Inst. 2002;94(13):972-980.
FREE FULL TEXT
9. Wei EK, Ma J, Pollak MN, et al.. A prospective study of C-peptide, insulin-like growth factor-I, insulin-like growth factor binding protein-1, and the risk of colorectal cancer in women. Cancer Epidemiol Biomarkers Prev. 2005;14(4):850-855.
FREE FULL TEXT
10. Vona-Davis L. Howard-McNattM, Rose DP. Adiposity, type 2 diabetes and the metabolic syndrome in breast cancer. Obes Rev. 2007;8(5):395-408.
PUBMED
11. Drew JE, Farquharson AJ, Padidar S, et al.. Insulin, leptin, and adiponectin receptors in colon: regulation relative to differing body adiposity independent of diet and in response to dimethylhydrazine. Am J Physiol Gastrointest Liver Physiol. 2007;293 (4):G682-G691.
FREE FULL TEXT
12. Williams CJ, Mitsiades N, Sozopoulos E, et al.. Adiponectin receptor expression is elevated in colorectal carcinomas but not in gastrointestinal stromal tumors. Endocr Relat Cancer. 2008;15(1):289-299.
FREE FULL TEXT
13. Fenton JI, Birmingham JM, Hursting SD, Hord NG. Adiponectin blocks multiple signaling cascades associated with leptin-induced cell proliferation in Apc Min/_ colon epithelial cells. Int J Cancer. 2008; 122(11):2437-2445.
PUBMED
14. Wei EK, Giovannucci E, Fuchs CS, Willett WC, Mantzoros CS. Low plasma adiponectin levels and risk of colorectal cancer in men: a prospective study. J Natl Cancer Inst. 2005;97(22):1688-1694.
FREE FULL TEXT
15. Filippi E, Sentinelli F, Romeo S, et al.. The adiponectin gene SNP_276G_T associates with earlyonset coronary artery disease and with lower levels of adiponectin in younger coronary artery disease patients (age _or=50 years). J Mol Med. 2005;83 (9):711-719.
PUBMED
16. Hara K, Boutin P, Mori Y, et al.. Genetic variation in the gene encoding adiponectin is associated with an increased risk of type 2 diabetes in the Japanese population. Diabetes. 2002;51(2):536-540.
FREE FULL TEXT
17. Heid IM, Wagner SA, Gohlke H, et al.. Genetic architecture of the APM1 gene and its influence on adiponectin plasma levels and parameters of the metabolic syndrome in 1,727 healthy Caucasians. Diabetes. 2006;55(2):375-384.
FREE FULL TEXT
18. Menzaghi C, Ercolino T, Di PR, et al.. A haplotype at the adiponectin locus is associated with obesity and other features of the insulin resistance syndrome. Diabetes. 2002;51(7):2306-2312.
FREE FULL TEXT
19. Qi L, Doria A, Manson JE, et al.. Adiponectin genetic variability, plasma adiponectin, and cardiovascular risk in patients with type 2 diabetes. Diabetes. 2006;55(5):1512-1516.
FREE FULL TEXT
20. Soccio T, Zhang YY, Bacci S, et al.. Common haplotypes at the adiponectin receptor 1 (ADIPOR1) locus are associated with increased risk of coronary artery disease in type 2 diabetes. Diabetes. 2006;55(10):2763-2770.
FREE FULL TEXT
21. Vasseur F, Helbecque N, Dina C, et al.. Singlenucleotide polymorphism haplotypes in the both proximal promoter and exon 3 of the APM1 gene modulate adipocyte-secreted adiponectin hormone levels and contribute to the genetic risk for type 2 diabetes in French Caucasians. Hum Mol Genet. 2002;11(21): 2607-2614.
FREE FULL TEXT
22. Crimmins NA, Martin LJ. Polymorphisms in adiponectin receptor genes ADIPOR1 and ADIPOR2 and insulin resistance. Obes Rev. 2007;8(5):419-423.
PUBMED
23. Mitchell MK, Gregersen PK, Johnson S, Parsons R, Vlahov D. The New York Cancer Project: rationale, organization, design, and baseline characteristics.J Urban Health. 2004;81(2):301-310.
PUBMED
24. Lukanova A, Soderberg S, Kaaks R, Jellum E, Stattin P.. Serum adiponectin is not associated with risk of colorectal cancer. Cancer Epidemiol Biomarkers Prev. 2006;15(2):401-402.
FREE FULL TEXT
25. Pollak MN, Schernhammer ES, Hankinson SE. Insulin-like growth factors and neoplasia. Nat Rev Cancer. 2004;4(7):505-518.
PUBMED
26. Moschos SJ, Mantzoros CS. The role of the IGF system in cancer: from basic to clinical studies and clinical applications. Oncology. 2002;63(4):317-332.
PUBMED
27. Ma J, Pollak M, Giovannucci E, et al.. A prospective study of plasma levels of insulin-like growth factor I (IGF-I) and IGF-binding protein-3, and colorectal cancer risk among men. Growth Horm IGF Res. 2000;10(suppl A):S28-S29.
PUBMED
28. Manousos O, Souglakos J, Bosetti C, et al.. IGF-I and IGF-II in relation to colorectal cancer. Int J Cancer. 1999;83(1):15-17.
PUBMED
29. Pischon T, Pai JK, Manson JE, et al.. Single nucleotide polymorphisms at the adiponectin locus and risk of coronary heart disease in men and women. Obesity (Silver Spring). 2007;15(8):2051-2060.
PUBMED
30. Kaklamani VG, Sadim M, Hsi A, et al.. Variants of the adiponectin and adiponectin receptor 1 genes and breast cancer risk. Cancer Res. 2008;68(9):3178-3184.
FREE FULL TEXT
31. Laken SJ, Petersen GM, Gruber SB, et al.. Familial colorectal cancer in Ashkenazim due to a hypermutable tract in APC. Nat Genet. 1997;17(1):79-83.
PUBMED
ARTICLE EN RAPPORT
Cette semaine dans le JAMA-Français
JAMA. 2008;300:1489.
Texte Complet
|