|
 
 |
|
ORIGINAL ARTICLE |
|
|
|
| Year : 2012 | Volume
: 18
| Issue : 2 | Page : 217-221 |
| |
The study of gene GJB2/DFNB1 causing deafness in humans by linkage analysis from district Peshawar
Zafar Ali1, Masroor Ellahi Babar1, Jamil Ahmad2, Sajjad Ali Shah1
1 Institute of Biochemistry and Biotechnology, University of Veterinary and Animal Sciences, Lahore, Pakistan
2 Faculty of Biotechnology and Informatics, Balochistan University of Information Technology, Engineering and Management Sciences, Queta, Pakistan
| Date of Web Publication | 8-Sep-2012 |
Correspondence Address:
Masroor Ellahi Babar
Director, Institute of Biochemistry and Biotechnology, University of Veterinary and Animal Sciences, Lahore
Pakistan
 Source of Support: None, Conflict of Interest: None
DOI: 10.4103/0971-6866.100771

 Abstract Abstract | | |
Families with at least 2 or more individuals having hereditary hearing loss were enrolled from different areas of Khyber Pakhtoonkhwa, mainly from district Peshawar. Detailed history was taken from each family to minimize the presence of other abnormalities and environmental causes for deafness. Families were questioned about skin pigmentation, hair pigmentation, and problems relating to balance, vision, night blindness, thyroid, kidneys, heart, and infectious diseases like meningitis, antibiotic usage, injury, and typhoid. The pedigree structures were based upon interviews with multiple family members, and pedigrees of the enrolled families were drawn using Cyrillic program (version 2.1). All families showed recessive mode of inheritance. I studied 8 families of these 10. For linkage analyses, studies for DFNB1 locus, 3 STR markers (D13S175, D13S292, and D13S787) were genotyped using polyacrylamide gel electrophoresis (PAGE) and haplotypes were constructed to determined, linkage with DFNB1 locus. From a total of 8 families, a single family-10 showed linkage to DFNB1 locus.
Keywords: Deafness, gene GJB2, linkage analysis, locus DFNB1
How to cite this article:
Ali Z, Babar ME, Ahmad J, Shah SA. The study of gene GJB2/DFNB1 causing deafness in humans by linkage analysis from district Peshawar. Indian J Hum Genet 2012;18:217-21 |
How to cite this URL:
Ali Z, Babar ME, Ahmad J, Shah SA. The study of gene GJB2/DFNB1 causing deafness in humans by linkage analysis from district Peshawar. Indian J Hum Genet [serial online] 2012 [cited 2016 Jun 1];18:217-21. Available from: http://www.ijhg.com/text.asp?2012/18/2/217/100771 |
| Introduction | |  |
Hearing is the process by which the ear transforms sound vibrations from the external environment into nerve impulses that are conveyed to the brain where they are interpreted as sounds. Hearing impairment is the partial or complete inability to hear that leads to compromise the development of normal language skills. Among all the sensory impairments in humans, hearing impairment is the most common. [1]
Profound hearing loss affects almost 1 in 1000 newborns, and more than 50% of these are caused by genetic factors. [2] It is estimated that the prevalence of profound bilateral hearing loss is 1.6 per 1000 in Pakistan, and 70% of hearing loss arises in consanguineous families. [3],[4] Hereditary hearing loss may be syndromic or non-syndromic; about 30% of deafness cases are syndromic, while 70% is non-syndromic. The main pattern of inheritance of deafness in Pakistani population is autosomal recessive. The common deafness syndromic is Usher, Pandred, and Waarenburg syndrome. The non-syndromic deafness may be autosomal dominant, autosomal recessive, X-linked, Y-linked, and mitochondrial. Approximately 75% cases of inherited deafness are autosomal recessive, 12-24% autosomal dominant, and 1-3% is X-linked. [5] To date, more than 67 loci and 21 genes have been identified for non-syndromic recessive deafness. GJB2 gene on chromosome 13q12-13 that codes for a gap junction protein called connexin 26 is responsible for the majority of genetic non-syndromic hearing losses. This transmembrane protein forms connexons in the cochlea that functions in potassium recycling in the hair cells.
Sound can be described in terms of frequency (pitch) measured in hertz (Hz), and in terms of intensity (loudness) measured in decibels (dB). Hearing is considered within the normal range if a person can process sound frequencies between 20 and 20,000 Hz and decibel levels from 0 to 140 dB. Hearing loss can be classified further by several criteria, including the severity (mild or 20 to 39 dB, moderate or 40 to 69 dB, severe or 70 to 89 dB, and profound or < 90 dB), age of onset (prelingual or postlingual), and the physiological etiology. Conductive hearing loss is characterized by external ear anomalies or abnormalities of the ossicles in the middle ear, sensorineural hearing loss is due to inner ear malfunction, and central hearing loss is caused by defects of the VIII th nerve, the brain stem, or the cerebral cortex. Hearing loss can also be mixed. [6],[7]
Connexin 26 (GJB2) is a member of gap junction proteins and can also interact with connexin 32, connexin 46, and connexin 50 to form a hexameric of homotypic (composed of six identical connexin subunits) or heterotypic (contain more than one species of connexins) half channel (connexon) of gap junctions. Such gap junction is responsible for the local transport of the potassium ions in the inner ear while other potassium channels pump potassium back into the endolymph [8] .
The major objective of this study was the linkage analysis for DFNB1 locus involved in causing hereditary deafness in some families from Khyber Pukhtoonkhwa. To achieve this objective, 10 families were enrolled through the schools for deaf children in Peshawar and surrounding areas. I studied 8 families i.e. family no. 2, 3, 4, 5, 6, 8, 9, and 10. Linkage analyses studies were carried out for DFNB1 locus. 3 STR markers D13S175, D13S787, and D13S292 were genotyped to determine if a family was linked to DFNB1 locus. A single family-10 showed linkage to DFNB1, and remaining 7 families were not linked to DFNB1 locus. GJB2 is a single exon gene that encodes connexin 26 protein.
It is anticipated that the present study will help to find out mutations responsible for deafness and will also make a contribution to future efforts in developing strategies for genetic counseling and prenatal diagnosis procedures for the population of Khyber Pukhtoonkhwa. This study will also lead to the discovery of new loci genes.
| Materials and Methods | | |
Enrollment of Families
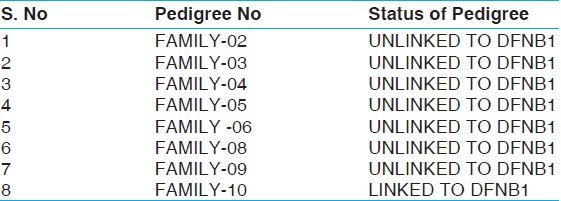
Families with at least 2 or more individuals having hereditary hearing loss were enrolled from different areas of Khyber Pakhtoonkhwa, mainly from district Peshawar. I enrolled a total of 10 families. Out of these 10 families, I studied 8 families i.e. 2, 3, 4, 5, 6, 8, 9, and 10. The status of these families are shown in the following [Table 1].
Deoxyribonucleic Acid Extraction
Blood samples of 5-10 ml were collected in 50 ml falcon tubes already containing 150 μl EDTA (0.5 M). Blood samples were frozen at −20°C for 24 hours before DNA extraction. DNA was extracted by inorganic method.
Washing of Blood
Washing of blood samples was processed with TE buffer (Tris HCl-10 mM, EDTA-2mM, pH = 8.0). It is also called lysis buffer. In this process, RBCs were broken down. Washing was done 3 or 4 times.
Digestion of Proteins
Proteinase K digestion was carried out by adding 45 μl of proteinase K along with 100 μl of 10% SDS in the presence of 3 ml buffer A-1 or TNE buffer (Tris HCL - 10 Mm, EDTA - 2 Mm, NaCl - 400 Mm), and the samples were digested over night in water bath shaker at 45°C. After digestion, the samples were ice-cold, and after that, left at room temperature for 10-15 minutes, followed by the addition of 1 ml of saturated 6 M NaCl. The samples were centrifuged, and DNA was extracted from the supernatant by adding equal volume of isopropanol. After washing with 70% ethanol, DNA was dissolved in TE buffer (10 Mm Tris, 0.2 Mm EDTA) and heated at 70°C for 1 hour to inactivate any remaining nucleases. Concentrations were kept at 25 ng/μl for single marker amplification.
Storage of the DNA
After that, the DNA was stored in screw tubes as a storage DNA solution. The DNA solution was transferred from falcon tubes to the screw tubes and was kept it in refrigerator to make its working solution.
Quantification of DNA
After extraction of DNA, the DNA quantification was done on 0.8% agarose gel.
Amplification of DNA
After that, PCR was carried out to amplify the DNA with the markers D13S175, D13S787, and D13S292.
Poly Acrylamide Gel Electrophoresis
The amplified PCR products were analyzed by using PAGE to show that if a family is linked or not to the locus DFNB1. The markers are shown in following [Table 2].
| Results | | |
A total of 10 families with multiple affected individuals were identified and enrolled from different areas of district Peshawar of Khyber Pakhtoonkhwa. I studied a total of 8 families. All the enrolled families were agreed to take part in the present study and to donate blood samples including affected and normal individuals. Blood samples (5-10 ml) were collected from affected individuals, normal siblings, and parents. During collection of blood samples, family members were interviewed for other syndromes in affected individuals. Pedigrees of the 8 families were drawn, using the Cyrillic program (version 2.1). After DNA extraction, linkage analysis studies were carried out. 3 STR markers D13S175, D13S292, and D13S787 were genotyped to determine if a family was linked or unlinked to DFNB1 locus. Hapalotypes (set of alleles) were constructed to determine the pattern of inheritance among the affected and normal individuals of each family under study. Out of 8 families which I was studied, a single family-10 was linked to DFNB1 locus.
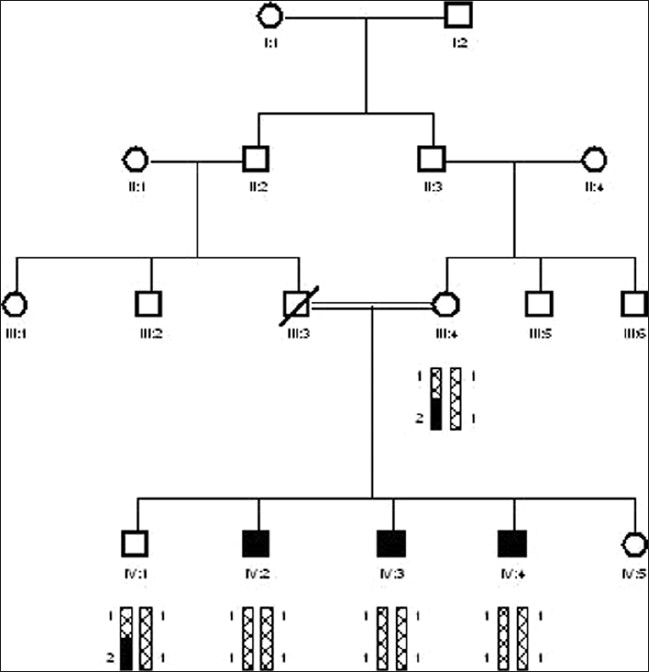
The family -10 was collected from Guli Khel FR Peshawar. Blood samples of 5 individuals were collected, having 3 affected (IV:2, IV:3, and IV:4) along with 1 normal sibling (IV:1) and mother (III:4). The affected individuals range in the age from 6, 12, and 16 years. All affected individual were appearing in 4th generation. After DNA isolation, 3 STR markers D13S175, D13S292, and D13S787 spanning in the region of DFNB1 were amplified. PCR product was electrophoresed on 6% polyacrylamide gel at 150 volts for 2 to 3 hours. Polyacrylamide gel was stained with ethedium bromide. Alleles were read manually with larger allele donated by 2 and smaller with 1. Haplotypes of family-10 showed linkage to BFNB1 locus. The affected individuals were homozygous with STR marker D13S292 and at DFNB1 candidate interval while normal sibling and mother were heterozygous .
| Discussion | | |
Non-syndromic deafness is a paradigm of genetic heterogeneity. Till date, more than 145 non-syndromic hearing loci (NSHL) have been genetically mapped; 58 of these loci are inherited in an autosomal dominant mode in which hearing loss is most frequently progressive and post-lingual in nature, of which only 21 genes have been identified. 8 non-syndromic deafness loci are x-linked, and 6 mutations of 2 mitchondrial genes are also indentified. The remaining 79 non-syndromic loci mapped so far are inherited as autosomal recessive, of which only 26 genes have been identified.
As a consequence of the unique socio-cultural practices in the population of Pakistan, approximately 60% of marriages are consanguineous, of which more than 80% are between first cousins. [9] Recessively inherited diseases are more prevalent in populations where cousin marriages are common, like Pakistan. [4] According to one estimate, the prevalence of profound bilateral hearing loss is 1.6 per 1000 in Pakistan, and 70% of hearing loss arises in consanguineous families. [3] These large consanguineous families are a powerful resource for genetic linkage studies of recessively inherited disorders like hearing impairment. [10]
Mutations in the GJB2 gene encoding connexin 26 are responsible for as much as 50% of pre-lingual, recessive deafness. By contrast, mutations in most of the other DFNB1 genes have so far been detected in only a small number of families, and their contribution to deafness on a population scale might, therefore, be limited. Identification of all genes involved in hereditary hearing loss will help in our understanding of the basic mechanisms underlying normal hearing, in early diagnosis and therapy. [11]
In this study, 10 families were selected for linkage analysis studies. These families were collected from different areas of Khyber Pakhtoonkhwa. I studied 8 families, which have at least 3 affected individuals. All the families have recessive mode of inheritance. Out of 8 families, a single family-10 showed linkage to DFNB1 locus [Figure 1]. The PAGE was done of all 8 families i.e. family no. 2, 3, 4, 5, 6, 8, 9, and 10 with all 3 markers i.e. D13S175, D13S787, and D13S292. They showed results with these markers. The Linked family has a total of 5 members with 3 affected individuals and normal mother and brother. It was collected from Guli Khel FR Peshawar.
Deafness at the DFNB1 locus is the most common cause of autosomal recessive non-syndromic deafness (ARNSD) in many countries throughout the world. [12] Typically caused by mutations in GJB2, the deafness is characteristically congenital and stable, varying in severity from moderate to profound. One GJB2 mutation, the 35delG allele variant, is the most common in populations of northern European ancestry. The carrier rate for this mutation among Caucasian population ranges from 1% to 3%. Mutations in the gene coding for connexin 26 account for about half of the autosomal recessive cases, corresponding to 10%-20% of all prelingual hearing impairment, and are, therefore, particularly important as a cause of genetic childhood hearing loss. One mutation, a deletion of a guanine residue at nucleotide 35 (35delG) resulting in a premature stop codon, has been found at high prevalence in most ethnic groups studied so far. The carrier frequency of this mutation has been estimated to be as high as 1/31 in people with a Mediterranean background. [13] The 35delG mutation causes a varied range of hearing losses, even among siblings or in individuals who are homozygous for this mutation. [7]
Mutations in the GJB2 gene are also responsible for syndromic forms of hearing loss, including autosomal dominant mutilating keratoderma with sensorineural deafness (Vohwinkel syndrome), other forms of autosomal dominant palmoplantar keratoderma with deafness, and the ectodermal dysplasia keratitis-ichtyosis-deafness syndrome. Most of these GJB2 mutations are located in the first extracellular domain. GJB2 is, therefore, an example of a connexin gene (like GJB3 and GJB6) associated with both non-syndromic deafness and genodermatosis with/without deafness. [11]
Further study is needed to explore the other mutations causing deafness in local population that will ultimately help to develop genetic counseling strategies and prenatal diagnostic procedures for the population of Khyber Pakhtoonkhwa.
| References | | |
| 1. | Agrup C, Gleeson M, Rudge P. The inner ear and the neurologist. J Neurol Neurosurg Psychiatry 2007;78:114- 22. 
[PUBMED] |
| 2. | Lalwani AK, Castelein CM. Nonsyndromic hereditary hearing impairment cracking the auditory genetic code. Am J Otol 1999;1:115-32.
|
| 3. | Elahi MM, Elahi F, Elahi A, Elahi SB. Pediatric hearing loss in rural are cause by allelic mutations of the novel cadherin-like gene CDH23. J Otolaryngol 1998;27:348-53.
[PUBMED] |
| 4. | Jaber L, Halpem GJ, Shohat M. The impact of consanguinity worldwide. Community Genet 1998;1:12-7.
|
| 5. | Marazita ML, Ploughman LM, Rawlings B, Remington E, Arnos KS, Nance WE. Genetic Epidemiological studies of early onset deafness in U.S school-age population. Am J Med Genet 1993;46:486-91.
[PUBMED] |
| 6. | Petit C, Levilliers J, Hardelin JP. Molecular genetics of hearing loss. Annu Rev Gene 2001;35:589-645.
[PUBMED] |
| 7. | Stephens D, Martini A, Mazzoli M. Audiological terms. Annu Rev Genet 2001;35:533-44.
|
| 8. | Xia JH, Liu CY, Tang BS, Pan Q, Huang L, Dai HP, et al. Mutations in the gene encoding gap junction protein beta-3 associated with autosomal dominant hearing impairment. Nat Genet 1998;20:370-3.
[PUBMED] |
| 9. | Hussain R, Bittles AH. The prevalence and demographic characteristics of consanguineous marriages in Pakistan. J Biosoc Sci 1998;30:261-75.
[PUBMED] |
| 10. | Khan SY, Ahmed ZM, Shabbir MI, Kitajiri S, Kalsoom S, Tasneem S, et al. Mutations of the RDX gene cause nonsyndromic hearing loss at the DFNB24 locus. Hum Mutat 2007;28:417-23.
[PUBMED] |
| 11. | Petersen MB, Willems PJ. Non-syndromic, autosomal-recessive deafness. Clin Genet 2006;69:371-92.
[PUBMED] |
| 12. | Zelante L, Gasparini P, Estivil X, Melchionda S, D'Agruma L, Govea N, et al. Connexin26 mutations associated with the most common form of non-syndromic neurosensory autosomal recessive deafness (DFNB1) in Mediterraneans. Hum Mol Genet 1997;6:1605-9.
|
| 13. | Estivill X, Govea N, Barcelo E, Badenas C, Romero E, Moral L, et al. Familial progressive sensorineural deafness is mainly due to the mtDNA A1555G mutation and is enhanced by treatment of aminoglycosides. Am J Hum Genet 1998;62:27-35.
|
[Figure 1]
[Table 1], [Table 2]
|