Targeting Cancer Stem Cells with Phytochemicals
Abstract
Cancer, second only to heart disease, is the leading cause of death in the US. Although progress has been made in the early detection of cancer and in improvements of cancer therapies, the ability to provide long-term survival has been limited. Increasing evidence suggests that a minute, biologically unique population of cancer stem cells (SCs) exists in most neoplasms and may be responsible for tumor initiation, progression, metastasis, and relapse. Characterization of cancer SCs has led to the identification of key cellular activities that may make cancer SCs vulnerable to therapeutic interventions that target drug-effluxing capabilities, stem cell pathways, anti-apoptotic mechanisms, and induction of differentiation. Phytochemicals, compounds made from fruits, vegetables, and grains, possess anti-cancer properties and represent a promising therapeutic approach for the prevention and treatment of many cancers. This review summarizes the evidence for the cancer SC hypothesis and discusses the potential mechanisms by which phytochemicals might target cancer SCs.

Introduction
Cancerous tumors are defined by uncontrolled proliferating cells that have lost their ability to be regulated by the host. Malignant tumors may remain localized for a time, but eventually they spread into the surrounding environment, enter into the circulatory system, and invade other tissues. Tumors comprise heterogeneous populations of cells that have varying degrees of tumorigenic potential. Only a subset of tumor cells are thought to initiate and promote tumorigenesis (1), and recent evidence has implicated a pluripotent subset of cells that have the capacity to seed the cellular heterogeneity seen in tumors (2). These cancer stem cells (SCs) are the earliest undifferentiated progenitors with an unlimited capacity to propagate. Given their potential role in tumorigenesis, cancer SCs are important targets for therapy. This review begins with a discussion of the cancer SC hypothesis and the relevance of these cells to tumorigenesis. We will then stress the importance of phytochemicals that show promise in attempts to target cancer SCs and biological processes relevant to drug effluxing capability, stem gene pathways, apoptotic pathways, and cellular differentiation (Table 1⇓).
Cancer Stem Cell Hypothesis
The cancer SC hypothesis proposes that a small subset of cells is responsible for the initiation, proliferation and metastasis of a tumor. Cancer SCs have the capacity to generate the heterogeneous population of cells that constitute a tumor (Figure 1⇓). Furthermore, these cells are resistant to a number of chemotherapeutic agents (3). In contrast, the bulk of the tumor (consisting of non-SCs) does not possess these qualities. From a clinical stand-point, the cancer SC hypothesis provides a compelling explanation for cancer patients who often relapse after treatment (radiation, surgery, or chemotherapy). Thus, if cancer SCs are resistant to therapy, then it may be the cancer SCs that promote the regrowth of tumor cells following withdrawal from chemotherapy (4, 5). It is of note that the name “cancer SC” does not necessarily imply its origins are from a “normal SC.” Cancer SCs may arise after mutations in early stem cell progenitors; however, it is equally possible that they are derived from mature, more differentiated cells. The term dedifferentiation, or transdifferentiation, has been used to describe this phenomenon (6). Thus, the operational term of a tumor- or cancer-initiating cell has also been used to avoid inferences of its source. Although beyond the scope of this review, the origins of cancer SCs continue to be under intense study (7–9).
Cancer SCs were first identified and chatacterized in patients with acute myeloid leukemia (AML) (10). Sorted cancer cell populations were divided into two groups (CD34+CD38– and CD34+CD38+ cells) and injected into nonobese diabetic/severe combined immunodeficiency (NOD/SCID) mice. The mice administered CD34+CD38– leukemic cells developed the disease whereas those given more mature cells (CD34+CD38−) did not. The leukemia formed in immunocompromised mice resembled the original patient disease in cellular morphology and in the ability of the cells to localize to the bone marrow. Further studies showed AML had a hierarchical organization that was similar to that seen in normal hematopoietic development (11), whereby early, multipotent progenitor cells give rise to later, more differentiated progeny cells. It is noteworthy that both leukemic and normal hematopoietic SCs shared similar cell-surface markers (CD34+CD38–) and both were able to recapitulate their respective systems. Additionally, serial transplantation of AML SCs resulted in the sustaining of the leukemic phenotype, and the morphology of these SCs was unchanged (11). As with hematopoietic SCs, the putative leukemic SCs were relatively quiescent and had the capacity to self-renew. Since the identification of leukemic SCs, cancer SCs have been found to reside in a number of other malignancies, including breast, brain, lung, prostate, colon, melanoma, and pancreatic cancers (12–18).
Properties of Cancer Stem Cells
Normal adult SCs contain both the ability to repopulate the cells that constitute the organ from which they are isolated and the capacity to propagate themselves. These processes must be tightly regulated. Cancer SCs reflect some of these same properties and are by defintion able to sustain proliferation; however, the cancer SC is not subject to the same genetic constraints to which a normal SC is bound. For example, many SC signaling components known to be involved in the maintenance and propagation of normal SCs are found to be mutated or aberrantly expressed in cancers, including Hedgehog (Hh), Wnt/β-catenin, Notch, Bmi-1, Hox, Oct3/4, and TGF-β (19, 20). Additionally, recent evidence suggests these SC molecules are important in regulating cancer SCs (21, 22). Liu et al. showed that Hh signaling and Bmi-1 expression are important regulators of breast cancer SC maintenance (23). A hallmark feature of breast cancer SCs is the formation of large, floating spheres (Box 1), termed mammospheres, that can be serially passaged (24). These mammospheres are highly tumorigenic and capable of forming colonies in vitro. Similar findings have been reported for brain tumor SCs (neurospheres) and prostate cancer SCs (prostatospheres) (14, 25). Treatment of mammospheres with soluble Hh showed 100% more secondary spheres and a 67% increase in the number of cells per sphere (23), suggesting Hh is an important regulator of SC self-renewal.
Spheres and Cancer SCs
Cancer SCs cells grow in round, unattached “spheres” in vitro. The first cancer SC spheres were identified and characterized in brain tumors (14). Purified CD133+ cells from primary human brain tumors were able to form “neurospheres,” whereas the CD133 cells could not. Cells were cultured in a chemically defined serum-free medium at a relatively low density (e.g., 1000 cells/well in a 96-well plate).
Cancer SCs have three unique properties that make them particularly important to tumor initiation, growth, and metastasis: self-renewal, multipotency, and a high proliferative capacity (5). Self-renewal is the ability of an SC to undergo symmetrical or asymmetrical division. Symmetrical division allows the cancer SCs to form either two differentiated daughter cells or two SCs. Asymmetric division results in one differentiated cell and one stem cell. The biological properties of self-renewal enables the SC to maintain its pluripotency while giving rise to differentiated progeny (26). Current methods of measuring self-renewal require the putative cancer SC to be serially transplantable (2–3 generations) into immunocompromised mice (4). For example, O’Brien et al. showed that primary colon cancer cells expressing CD133 were the putative cancer SCs (16). The CD133+ cells formed tumors in NOD/SCID mice, whereas the CD133– cells (which comprised the bulk of the tumor) did not. CD133+ cells were subsequently isolated from the parental mouse tumor and serially transplanted back into NOD/SCID mice. Secondary and tertiary transplantation of the CD133+ cells continued to form tumors, demonstrating self-renewal capacity of the cancer SCs. Each tumor that formed from the serial transplantation appeared histologically similar to the parental mouse tumor.
Multipotency is the ability of cancer SCs to differentiate into the heterogeneous population of cells that form the tumor. This can be demonstrated by inducing putative cancer SCs toward a more differentiated cell fate. For example, Hurt et al. showed that LNCaP prostate cancer cells expressing CD44+CD24– behaved as prostate cancer SCs (25). These cells were highly tumorigenic and expressed a number of genes important for SC self-renewal and maintenance, including Oct-3/4 and Bmi-1. The putative cancer SCs were cultured in a serum replacement medium and formed large, floating prostato-spheres. When the cells were exposed to serum, they adhered to the culture plate and exhibited decreased expression of the stem cell genes. Moreover, gene expression profiling showed that the serum-treated cancer SCs genotypically resembled the parental cell line, suggesting the serum-treated prostate cancer SCs differentiated toward the heterogeneous cell population.
Before the discovery of the cancer SC, injection of millions of tumor cells was required to form tumors in rodent models; however, as little as one hundred cancer SCs are needed to form tumors (4). For example, Al-Hajj et al. isolated primary breast cancer SCs that were CD44+CD24–/loESA+ (12); these tumor-initiating cells formed a tumor in NOD/SCID mice with as few as 100 cells, whereas their depleted counterparts required 200-fold more.
The existence of cancer SCs and their ability to self-renew, differentiate into multiple lineages, and proliferate extensively make them particularly insidious. If for a given tumor, a cancer SC is responsible for disease manifestation, future therapies that are not effective against cancer SCs may fall short of providing long-term disease management.
Targeting Cancer Stem Cells Using Phytochemicals
The ability of phytochemicals to inhibit tumor formation both in vitro and in vivo is well documented (27). Many of these compounds have anti-oxidant, anti-proliferative, and pro-apoptotic effects on a variety of cancers, including leukemia, prostate, breast, colon, brain, melanoma, and pancreatic (27–30). Moreover, cancer SCs have been identified for each of these blood and solid tumors (10, 12, 13, 16, 17, 20, 31, 32), paving the way for future studies to evaluate the efficacy of phytochemicals against cancer SCs. The benefit of many of these phytochemicals is that they are well tolerated and are found in many food products that can be added to one’s diet (33). Furthermore, phytochemicals could be taken on a long-term basis to either prevent primary tumor formation or tumor recurrence (33).
An extensive amount of research is being conducted on the benefits of phytochemicals. Indeed, there are a number of clinical trials that have evaluated these natural products for their efficacy in preventing disease (Table 2⇓). For example, a mixture of green tea catechins was used in a chemoprevention trial on patients with high-grade prostate intraepithelial neoplasia (HG-PIN), the main premalignant lesion of prostate cancer (34). Preclinical data had shown that green tea catechins, particularly (−)-epigallocatechin-3-gallate (EGCG), have anti-cancer activities (35). Patients were given three daily treatments (600 mg/day) of the mixed catechins. After one year, the conversion rate from HG-PIN to prostate cancer was 3% in the experimental group and 30% in the placebo group. Another clinical trial looked at the effects of vitamin D on patients with progressive, metastatic androgen-independent prostate cancer. Vitamin D and its analogs are anti-proliferative and anti-angiogenic in a wide range of cancers (36). The study found that vitamin D in combination with the standard chemotherapy agent, docetaxel, led to an improvement in overall patient survival (37). Further confirmation in this study is required, however, because survival was not the primary endpoint. Taken together, the results emerging from clinical data suggest phytochemicals will be valuable tools in the prevention or treatment of cancer.
Targeting Drug Pumps
Although chemotherapy can reduce tumor mass, an aggressive population of cancer SCs within the tumor may be capable of resisting chemotherapeutic drugs (3), leading to relapse and mul-tidrug resistance (MDR) (38). For example, cancer SCs from brain tumors express the neural stem cell surface marker CD133+ (14). Cancer SCs isolated from patient glioblastomas were more resistant to chemotherapeutic agents (e.g., temozolomide, carboplatin, paclitaxel, and etoposide) than were their non-cancer SC counterparts (CD133–). Additionally, CD133 expression was higher in recurrent glioblastomas as compared to that in a newly diagnosed patient, suggesting the CD133+ cancer SCs were better able to survive therapy. Finally, Shervington and Lu found a close association between CD133 and MDR expression in glioblastoma tissue (40). Greater MDR expression has also been associated with cancer SCs from melanomas, pancreas, and breast (41–43).
Natural Compounds that Target Multidrug Resistance
The major mechanism of drug efflux in MDR is carried out by energy-dependent transporter proteins belonging to the ATP-binding cassette (ABC) family. Members of this group of transporter proteins include P-glycoprotein (P-gp), multidrug resistant-associated proteins (MRPs), and mitoxantrone resistance protein (MXR or ABCG2). Although clinical trials have yet to assess the role of natural products against MDR, a variety of natural products interact with ABC transport proteins and reverse the MDR phenotype in vitro. Specifically, plant polyphenols and the phytochemical curcumin (found in the spice turmeric) have been extensively studied.
Polyphenol compounds are active agents in green tea and include EGCG and epigallocatechin (EGC), the most biologically active catechins. In Chinese hamster ovary (P-gp+) cells, green tea–derived polyphenols inhibited P-gp transport activities, as evidenced by the accumulation of rhodamine-123 dye within the cells (44). The anti-cancer effects of EGCG were enhanced when combined with vinblastine (an anti-mitotic drug) or doxorubicin (a DNA-interacting anthracycline antibiotic), suggesting EGCG facilitated a reduction in drug efflux and a reversal of the MDR phenotype (45–47). Wu et al. demonstrated that certain polyphenols (i.e., quercetin, silymarin, resveratrol, naringenin, daidzein, and hesperetin) inhibited the MRP1, 4, and 5 ABC transporters (48). The investigators measured efflux inhibition using fluorescent substrate or radiolabeled-vinblastine accumulation in MRP1-expressing HEK293 cells.
Curcumin is the most potent polyphenol in turmeric, a spice widely consumed in Southeast Asian countries. Combination treatment of curcumin with a variety of chemotherapy drugs (i.e., tamoxifen, cisplatin, danorubicin, doxorubicin, and vincristine) enhances the cellular accumulation of these drugs (49, 50), effectively sensitizing drug-resistant cells. Choi et al. treated L1210/ Adr cells (a multidrug-resistant mouse leukemia cell line) with curcumin and observed decreased expression of mdr1b, mediated by the phosphoinositide-3′ kinase (PI3K), Akt, and nuclear factor-kappaB (NF-κB) pathways (51). Curcumin also inhibited the activity of the transporter ABCG2 (52). Addition of curcumin reversed MDR in ABCG2-expressing HEK cells and facilitated the accumulation of doxorubicin and mitoxantrone in cells, thereby increasing the cells sensitivity to the chemotherapeutic drugs.
Cancer SCs may exploit MDR to lower the cellular accumulation of chemotherapeutic drugs and thereby sustain tumor growth. Natural products show great promise as anti-MDR agents, and given the strong link between cancer SCs and MDR, may reflect the feasibility of targeting cancer SCs to fight dise (3).
Targeting Stem Cell Signaling Pathways: Hedgehog, Wnt, Notch
Little is known about the molecular mechanisms that control self-renewal of SCs, an essential element of tumor survival and propagation (23, 53, 54). Genes expressed in embryonic SCs, encoding proteins involved in Hedgehog (Hh), Wnt/β-catenin, and Notch signaling, are key factors in regulating self-renewal. Although these genes are expressed in normal SCs, they are frequently mutated or aberrantly activated in cancers, thus making them potential therapeutic targets.
Hedgehog
Hh is a secreted protein involved in a number of vertebrate developmental processes, including polarity in the central nervous system, left-right asymmetry, organogenesis and spermatogenesis (55). Hh signaling proceeds through two transmembrane proteins, Patched (PTC), and Smoothened (Smoh). At rest, PTC inhibits Smoh activity, but upon its binding to Hh, releases this inhibition, thus allowing Smoh-mediated activation followed by Hh related transcriptional responses (56). Perturbations in the Hh pathway are linked to cancer SCs in glioblastomas, multiple myeloma, pancreas, and breast (23, 57–59).
The first phytochemical shown to inhibit the Hh pathway was cyclopamine (60), a naturally occurring compound found in the plant Veratrum californicum, commonly called the corn lily. Cyclopamine targets the Hh pathway, specifically by inhibiting the activation of Smoh (61). Cyclopamine treatment of a murine medulloblastoma blocked proliferation and induced neuronal differentiation, effectively depleting the cancer SC population (62). This study further demonstrated that cyclopamine reduced tumor burden in a mouse tumor allograft and was cytotoxic to cultured human medulloblastoma cells. In addition, cyclopamine is effective in targeting cancer SCs of pancreatic cancer, breast cancer, and multiple myeloma (23, 58, 59). Sonic Hh (Shh) expression is elevated in pancreatic cancer SCs (17), the putative mediators of pancreatic tumor invasion and metastasis (32). Combination therapy of cyclopamine and gemcitabine inhibited metastatic spread and reduced primary tumor burden in pancreatic orthotopic xenografts (59). In breast cancer and multiple myeloma SCs, cyclopamine reduced mammosphere formation and SC proliferation, respectively (23, 58). Taken together, cyclopamine targets the self-renewal properties of cancer SCs, highlighting an important exploitable weakness.
Wnt/β-Catenin
Wnt/β-catenin signaling is involved in a wide range of developmental processes including maintenance of stem cell compartments in adult tissue (63). Wnt is a protein tethered to the extracellular matrix that activates the low-density lipoprotein receptor–related protein (LRP) and receptors of the Frizzled family of proteins. In the absence of Wnt, a complex of at least three proteins––cytoplasmic enzyme glycogen synthase kinase-3β (GSK-3β), axin, and adenomatous polyposis coli (APC)––destabilizes β-catenin and targets it for destruction. Activation of Wnt stabilizes β-catenin, which acts as a transcriptional coactivator for a number of developmental responses (64). Mutations in the Wnt/β-catenin pathway are associated with a number of cancers (65) and implicated in controlling cancer SC self-renewal capabilities (66). Several phytochemicals, such as selenium, EGCG, and vitamin D, were recently shown to inhibit Wnt signaling in cancers and could potentially be excellent candidates for targeting cancer SCs (67–69).
Selenium is an essential trace element found in a variety of compounds, including inorganic salts, amino acids, and methylated compounds. Research dating back to the 1960s suggested selenium may have anti-carcinogenic properties (70). A study done on colon cancer cells showed that an organoselenium compound, 1,4-phenylene bis(methylene) selenocyanate (p-XSC), in combination with docosahexaenoic acid (DHA), an ω-3 fatty acid, synergistically reduced the viability of colon cancer cells with a concomitant reduction in β-catenin expression (71). The same p-XSC compound decreased polyp formation in ApcMin/+ mice, an animal model for familial adenomatous polyposis (FAP) (67). The ApcMin/+ mouse has an autosomal dominant heterozygous nonsense mutation of the mouse Apc gene and exhibit spontaneous gastrointestinal tumors similarly found in humans with germ line and somatic Apc mutations (72). In addition, treatment with p-XSC also led to reduced expression of β-catenin in the adenomas.
EGCG altered Wnt/β-catenin signaling in breast cancer cells (68). Kim et al. showed EGCG inhibited Wnt-induced gene expression responses such as reduced activity of TCF/LEF binding and decreased c-Myc expression. This attenuation of Wnt/β-catenin activity was mediated through the stabilization of HBP-1, a transcriptional repressor of Wnt/β-catenin signaling and a suppressor of oncogenesis (65). EGCG also inhibited tumor formation in ApcMin/+ mice (73). The investigators determined that nuclear β-catenin levels were significantly reduced, further implicating the Wnt/β-catenin signaling pathway.
Vitamin D can also mediate some of its anti-cancer effects through inhibition of Wnt signaling. In colon cancer, vitamin D inhibited the expression of β-catenin–responsive genes, thereby reducing levels of c-myc, peroxisome proliferator-activated receptor, Tcf-1, and CD44 (74). Vitamin D altered β-catenin subcellular localization from predominantly nuclear to almost exclusively cytoplasmic. In addition, it induced expression of E-cadherin, a scaffolding and tumor-suppressing protein involved in sequestering and inhibiting the activity of β-catenin. Additionally, overex-pression of the vitamin D receptor counteracted the inhibitory effects of vitamin D upon Wnt/β-catenin signaling (75).
Notch
Notch signaling is activated by direct cell–cell contact. Activation of Notch leads to proteolytic cleavage of the intracellular domain of Notch (ICN). The ICN cleavage product then translocates to the nucleus and coordinates a transcription factor complex that includes CSL (an acronym for vertebrate CBF/RBP-jk, Drosophila Su(H), and C. elegans Lag-1) and co-activators of the Mastermind-like family of proteins (76). Notch and its signaling components are implicated in a wide variety of developmental processes, such as central nervous system development, vasculature system development, organogenesis, and adult-type hematopoietic stem cell generation (76). In addition, Notch signaling promotes the expansion of neuronal and breast SCs (77, 78). Aberrant Notch signaling is implicated in controlling tumor self-renewal in medulloblastomas (79). Notch expression was found to be enriched in brain tumor SCs, and inhibition of Notch signaling resulted in apoptosis in the cancer SC population and blocked xenograft tumor formation.
The phytochemical resveratrol (found in grapes, berries and peanuts) exhibits cardioprotective and anti-cancer properties (80, 81). Acute lymphoblastic leukemia cells treated with resveratrol resulted in decreased Notch protein expression (82). Resveratrol appeared to affect Notch at the post-translational level because Notch mRNA levels were not affected. Moreover, mRNA levels of downstream effectors of Notch [e.g., pre-TCRα and Hairy and Enhancer of Split (HES1)] were decreased in the presence of resveratrol. In addition to Notch signaling, resveratrol also inhibited the Akt-PI3K pathway and activated the proapoptotic/tumor suppressor p53 signaling pathway.
Targeting Apoptosis (NF-κB)
The NF-κB pathway is involved in a tumor’s ability to proliferate, evade apoptosis, invade, and metastasize (83). NF-κB consists of homo- and heterodimers sequestered in the cytoplasm by the protein inhibitor of NF-κB (IκB) (84). The NF-κB signaling pathway is activated by bacterial and viral infections, oxidative stresses, and growth factors or cytokines, and results from the translocation of NF-κB into the nucleus, where it regulates gene expression (83). Guzman et al. demonstrated that NF-κB is aberrantly expressed in leukemic SCs but not in normal primitive hematopoietic cells (85). Moreover, treatment of the leukemic SCs with a potent NF-κB inhibitor rapidly induced cell death, suggesting the NF-κB pathway may be a therapeutic target.
Parthenolide is a naturally occurring compound found in feverfew, a plant whose extracts have been used in folk remedies to control fever and headache severity. Parthenolide has been shown to target acute myelogenous leukemia (AML) SCs and their progenitors while sparing normal hematopoietic cells (86). The molecular mechanism of parthenolide action was associated with inhibiting NF-κB–mediated apoptosis, activation of the p53 proapoptotic pathway, and increased reactive oxygen species. In addition to AML, parthenolide targeted mammary breast cancer SCs (43). In vitro, parthenolide preferentially inhibited mammosphere growth for greater than seventy-two hours (43). The decrease of sphere growth was due to the inhibition of NF-κB activity.
Differentiation of Cancer Stem Cells
A primary characteristic of cancer SCs is their ability to differentiate into heterogeneous cells with differing rates of proliferation. Until recently, most cancers were treated using anti-proliferation agents that affected cells at various points in the cell cycle (87). Several treatment cycles are required to ensure that tumor cells at all stages of the cell cycle are affected. Despite many rounds of chemotherapy, the patient often relapses, suggesting this approach does not effectively target the quiescent cancer SC (88). Therefore, inducing cancer SCs to differentiate into proliferative stages that would be labile to chemotherapeutics may be an effective step in combatting cancer.
Vitamin A is a fat-soluble nutrient found in a number of natural products including carrots, broccoli, and liver. Retinoic acid (RA) is the active form of vitamin A found in animals, whereas β-carotene is a precursor to the active form of vitamin A in plants (89). RAs play a critical role in embryogenesis and hemotopoiesis (90, 91) and RA has thus been seen as a potential mediator of differentiation therapy. Proof-of-principle was achieved with the treatment of acute promyelocytic leukemia (APL) (92). APL is caused by a chromosomal translocation that produces a chimeric protein between the retinoic acid receptor-alpha (RARα) and a protein called promyelocyte leukemia protein (PML). Addition of retinoic acid induces differentiation of promyelocytic leukemic cells to mature neutrophils (93). As an adjunct to clinical therapy, RA dramatically improved patient outcome, with complete remission rates of greater than 90% and long-term survival in about 75% of patients (94).
Concluding Remarks
Phytochemicals are natural products found in our diet and can be used to target cancer SCs. However, phytochemicals are not without their disadvantages. For example, the wide use of St. John’s wort (Hypericum perforatum) may have detrimental effects in anti-cancer therapies (95). The phytochemical in the wort can alter drug pharmacokinetics by modulating the activity of P-gp and CYP3A4, an enzyme that oxidizes many anti-cancer drugs (95). The net result is a reduction in oral absorption and elimination of the therapeutic drug. Given the clinical implications and the widespread use of these phytochemicals, rigorous testing for possible adverse interactions are required as well as stringent quality control protocols. Even with these caveats, however, phytochemicals still represent a promising addition to standard therapy.
The advantages of phytochemicals are: 1) they may potentially help to prevent the occurrence of cancer by eating a healthy diet rich in anti-cancer phytochemicals; 2) certain phytochemicals show promise as a complement to current chemotherapy—for example, vitamin D combined with conventional therapy is already in clinical trials to treat prostate and breast cancer (36); and 3) phytochemicals may be used over time after conventional therapy, ostensibly to keep cancer SCs in check and limit relapse. Investigating the properties of phytochemicals and their role in targeting cancer SCs will further enhance our understanding of how cancer SCs contribute to a growing tumor and enable us to develop treatment strategies.
Targeting Cancer Stem Cell Properties
Evaluation of Phytochemicals in Clinical Trials
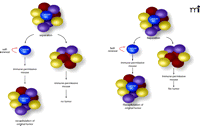
The cancer stem cell hypothesis. Only the cancer SC is capable of initiating tumor growth and giving rise to the diverse population of cancer cells (represented by different colored cells). Cancer SCs have the ability to self-renew (symmetrical and asymmetrical division) and demonstrate multipotency. Additionally, cancer SCs must be able to recapitulate the heterogeneity of the original tumor.
Tashan Mistree, MS, (photo unavailable) obtained her undergraduate degree from Georgia Institute of Technology and subsequently received a Master’s degree from Johns Hopkins University.
Acknowledgments
We would like to thank members of the Farrar laboratory for their helpful discussions and editing contributions, especially George Klarmann. This research was supported in part by the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research. In addition, this work was funded in part by the Office of Dietary Supplements, National Institutes of Health. The content of this paper does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the US Government.
- © American Society for Pharmacology and Experimental Theraputics 2008
References

William L. Farrar, PhD, obtained his degree from the Virginia Polytechnic Institute. His postdoctoral work focused on cytokines and their signaling. Currently, he is the head of the Cancer Stem Cells Section in the Laboratory of Cancer Prevention at the National Cancer Institute in Frederick, MD where his studies are now focused on understanding the underlying biology of cancer stem cells. Using an integrated molecular profiling approach, he and his colleagues are investigating the mechanisms underpinning cancer stem cell differentiation, self-renewal, and invasion. E-mail: far-rar{at}mail.ncifcrf.gov; fax: 301-846-6104.1

Elaine M. Hurt, PhD, obtained her degree from the University of Minnesota where she studied estrogen receptor transcriptional responses. Subsequent postdoctoral work in the laboratory of Louis M. Staudt, MD, PhD, at the National Cancer Institute in Bethesda focused on gene expression profiling of lymphoma and multiple myeloma as well as understanding the role of c-maf in the etiology of myeloma. Currently, she is a staff scientist in the laboratory of Dr. Farrar where her studies are concentrated on understanding self-renewal and differentiation of prostate cancer stem cells. E-mail: hurte{at}ncifcrf.gov; fax: 301-846-6104.

Brian T. Kawasaki, PhD, obtained his degree from the University of California, Los Angeles where he studied the role of the tyrosine kinase Src in regulating capacitative calcium entry channels. Subsequent post-doctoral work in the laboratory of Dr. William L. Farrar at the National Cancer Institute in Frederick, MD focused on using natural products to target prostate cancer stem cells. Dr. Kawasaki has shifted careers and is currently enrolled in medical school at the University of Maryland, School of Medicine.

